 墍揷偝傫偼嬨廈戝妛偺堛妛晹傪懖嬈偟丄撪壢偱摐擜昦偺愱栧堛偲偟偰俀擭娫尋廋偟傑偟偨丅丂偦偺屻丄戝妛傊栠偭偰戝妛堾偱尋媶傪偡傞帪婜偵側傝傑偟偨偑丄柤榓揷嫵庼偺悇慐偱嫗戝偺巹偨偪偺尋媶幒偵戝妛堾惗偲偟偰庴尡偟丄杮奿揑側尋媶妶摦傪奐巒偡傞偙偲偵側傝傑偟偨丅丂偛傜傫偺捠傝丄偦偟偰丄僶僗働僢僩偵僾儘偲偄偆丄僫僀僗僈僀丅丂偱傕丄偪傖傫偲嫗搒傊棃傞慜偵丄惍宍奜壢堛偺墱偝傫傪僎僢僩偟偰偄傑偡丅丂戝妛堾偵擖偭偰俀擭栚偱偙偺榑暥傪巇忋偘傑偟偨丅丂僋儘乕僨傿儞偺婡擻夝愅偲偟偰偼弶偺榑暥偱偡丅
墍揷偝傫偼嬨廈戝妛偺堛妛晹傪懖嬈偟丄撪壢偱摐擜昦偺愱栧堛偲偟偰俀擭娫尋廋偟傑偟偨丅丂偦偺屻丄戝妛傊栠偭偰戝妛堾偱尋媶傪偡傞帪婜偵側傝傑偟偨偑丄柤榓揷嫵庼偺悇慐偱嫗戝偺巹偨偪偺尋媶幒偵戝妛堾惗偲偟偰庴尡偟丄杮奿揑側尋媶妶摦傪奐巒偡傞偙偲偵側傝傑偟偨丅丂偛傜傫偺捠傝丄偦偟偰丄僶僗働僢僩偵僾儘偲偄偆丄僫僀僗僈僀丅丂偱傕丄偪傖傫偲嫗搒傊棃傞慜偵丄惍宍奜壢堛偺墱偝傫傪僎僢僩偟偰偄傑偡丅丂戝妛堾偵擖偭偰俀擭栚偱偙偺榑暥傪巇忋偘傑偟偨丅丂僋儘乕僨傿儞偺婡擻夝愅偲偟偰偼弶偺榑暥偱偡丅Sonoda,N., Furuse,M., Sasaki,H., Yonemura,S., Katahira,J., Horiguchi,Y., and Tsukita,Sh.丂
J.Cell Biol. 丂147,195-2041 (1999)
丂
亂昅摢挊幰徯夘亃
墍揷偝傫偼嬨廈戝妛偺堛妛晹傪懖嬈偟丄撪壢偱摐擜昦偺愱栧堛偲偟偰俀擭娫尋廋偟傑偟偨丅丂偦偺屻丄戝妛傊栠偭偰戝妛堾偱尋媶傪偡傞帪婜偵側傝傑偟偨偑丄柤榓揷嫵庼偺悇慐偱嫗戝偺巹偨偪偺尋媶幒偵戝妛堾惗偲偟偰庴尡偟丄杮奿揑側尋媶妶摦傪奐巒偡傞偙偲偵側傝傑偟偨丅丂偛傜傫偺捠傝丄偦偟偰丄僶僗働僢僩偵僾儘偲偄偆丄僫僀僗僈僀丅丂偱傕丄偪傖傫偲嫗搒傊棃傞慜偵丄惍宍奜壢堛偺墱偝傫傪僎僢僩偟偰偄傑偡丅丂戝妛堾偵擖偭偰俀擭栚偱偙偺榑暥傪巇忋偘傑偟偨丅丂僋儘乕僨傿儞偺婡擻夝愅偲偟偰偼弶偺榑暥偱偡丅
 Clostridium perfringens偲偄偆嵟嬤偼儁僾僠僪惈偺挵撪撆慺乮俠俹俤乯傪暘斿偟丄寉偄怘拞撆傪婲偙偡丅丂嶃戝旝尋偺曅暯傜偼丄偙偺撆慺偺嵶朎懁偺儗僙僾僞乕傪摨掕偟丄俠俹俤儗僙僾僞乕乮俠俹俤亅俼乯偲曬崘偟偰偄偨丅丂変乆偑僋儘乕僨傿儞乕侾丆乕俀傪摨掕偟偰傒傞偲丄偙偺俠俹俤亅俼偵帡偰偄傞偙偲偑暘偐傝丄俠俹俤亅俼傪僋儘乕僨傿儞乕係偲屇傇傛偆偵側偭偨丅丂偡側傢偪丄偙偺撆慺偼僋儘乕僨傿儞乕係傪儗僙僾僞乕偲偟偰棙梡偟偰偄偨偙偲偵側傞丅丂偙偺撆慺偼丄偦偺俠枛抂敿暘偱僋儘乕僨傿儞乕係乮俠俹俤乕俼乯偵寢崌偟丄俶枛抂敿暘偱嵶朎枌偵寠傪偁偗傞偙偲偑暘偐偭偰偄偨丅丂偦偙偱丄偙偺撆慺偺俠枛抂敿暘偺儁僾僠僪乮俠亅俠俹俤乯偼丄僋儘乕僨傿儞乕係偵摿堎揑偵寢崌偡傞儁僾僠僪偲偟偰棙梡偱偒傞偺偱偼側偄偐偲峫偊傞傛偆偵側偭偨丅
Clostridium perfringens偲偄偆嵟嬤偼儁僾僠僪惈偺挵撪撆慺乮俠俹俤乯傪暘斿偟丄寉偄怘拞撆傪婲偙偡丅丂嶃戝旝尋偺曅暯傜偼丄偙偺撆慺偺嵶朎懁偺儗僙僾僞乕傪摨掕偟丄俠俹俤儗僙僾僞乕乮俠俹俤亅俼乯偲曬崘偟偰偄偨丅丂変乆偑僋儘乕僨傿儞乕侾丆乕俀傪摨掕偟偰傒傞偲丄偙偺俠俹俤亅俼偵帡偰偄傞偙偲偑暘偐傝丄俠俹俤亅俼傪僋儘乕僨傿儞乕係偲屇傇傛偆偵側偭偨丅丂偡側傢偪丄偙偺撆慺偼僋儘乕僨傿儞乕係傪儗僙僾僞乕偲偟偰棙梡偟偰偄偨偙偲偵側傞丅丂偙偺撆慺偼丄偦偺俠枛抂敿暘偱僋儘乕僨傿儞乕係乮俠俹俤乕俼乯偵寢崌偟丄俶枛抂敿暘偱嵶朎枌偵寠傪偁偗傞偙偲偑暘偐偭偰偄偨丅丂偦偙偱丄偙偺撆慺偺俠枛抂敿暘偺儁僾僠僪乮俠亅俠俹俤乯偼丄僋儘乕僨傿儞乕係偵摿堎揑偵寢崌偡傞儁僾僠僪偲偟偰棙梡偱偒傞偺偱偼側偄偐偲峫偊傞傛偆偵側偭偨丅
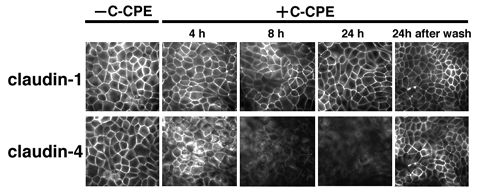
搒崌偺椙偄偙偲偵丄忋恾偺傛偆偵丄攟梴俵俢俠俲忋旂嵶朎偼丄庡偵僋儘乕僨傿儞乕侾偲乕係傪敪尰偟偰偄傞丅丂偦偙偱丄偙偺嵶朎傪俠亅俠俹俤懚嵼壓偱攟梴偟偰傒偨丅丂偡傞偲丄偐側傝憗偄帪娫宱夁偱僞僀僩僕儍儞僋僋僔儑儞偐傜丄僋儘乕僨傿儞乕係偩偗偑徚幐偟丄僋儘乕僨傿儞乕侾偩偗偑巆傞偙偲偑暘偐偭偨丅丂偙偺俠亅俠俹俤傪愻偄棳偡偲僋儘乕僨傿儞乕係偼僞僀僩僕儍儞僋僔儑儞偵栠偭偰偔傞丅
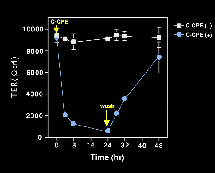 偙偺夁掱偱忋旂嵶朎偺僶儕傾乕婡擻偑偳偆曄傢傞偐傪俿俤俼傪寁傞偙偲偵傛傝應掕偟偨偺偑嵍恾偱偁傞丅丂僋儘乕僨傿儞乕係偺徚幐偺帪娫宱夁偵堦抳偟偰僶儕傾乕婡擻偑嬌抂偵掅壓偡傞偙偲偑暘偐偭偨丅
偙偺夁掱偱忋旂嵶朎偺僶儕傾乕婡擻偑偳偆曄傢傞偐傪俿俤俼傪寁傞偙偲偵傛傝應掕偟偨偺偑嵍恾偱偁傞丅丂僋儘乕僨傿儞乕係偺徚幐偺帪娫宱夁偵堦抳偟偰僶儕傾乕婡擻偑嬌抂偵掅壓偡傞偙偲偑暘偐偭偨丅
 偙偺帪僞僀僩僕儍儞僋僔儑儞偺峔憿偼偳偆側偭偰偄傞偱偁傠偆偐丅丂嵍恾偺傛偆偵丄僶儕傾乕婡擻偑幐傢傟傞偺偲摨帪偵丄僞僀僩僕儍儞僋僔儑儞偺僗僩儔儞僪偑梟偗巒傔丄嵟廔揑偵偼偒傢傔偰昻庛側僱僢僩儚乕僋偵曄壔偡傞偙偲偑暘偐偭偨丅
偙偺帪僞僀僩僕儍儞僋僔儑儞偺峔憿偼偳偆側偭偰偄傞偱偁傠偆偐丅丂嵍恾偺傛偆偵丄僶儕傾乕婡擻偑幐傢傟傞偺偲摨帪偵丄僞僀僩僕儍儞僋僔儑儞偺僗僩儔儞僪偑梟偗巒傔丄嵟廔揑偵偼偒傢傔偰昻庛側僱僢僩儚乕僋偵曄壔偡傞偙偲偑暘偐偭偨丅
埲忋偺寢壥偼丄僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪偺僋儘乕僨傿儞偵傛傞宍惉偑丄忋旂嵶朎偺僶儕傾乕婡擻偦偺傕偺傪扴偭偰偄傞偙偲傪偼偠傔偰幚尡揑偵帵偟偨傕偺偱偁傝丄嵍偺傛偆偵俰俠俛偺昞巻傪忺傞偙偲偵側偭偨丅
![]()