 椺偊偽丄嵶朎幙暘楐偺帪偵丄嵶朎偺愒摴柺偺嵶朎枌捈壓偵傾僋僠儞僼傿儔儊儞僩偑廤傑傝丄嵶朎枌傪撪懁傊堷偒崬傓偙偲偵傛傝暘楐峚偑偱偒傞偺偼桳柤偱偡丅丂偟偐偟丄傾僋僠儞僼傿儔儊儞僩偲嵶朎枌偑偳偺傛偆側暘巕婡峔偱寢崌偟偰偄傞偐偼丄撲偺傑傑偱偟偨丅
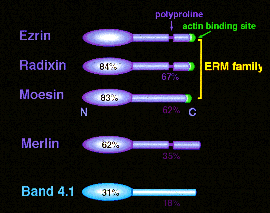
椺偊偽丄嵶朎幙暘楐偺帪偵丄嵶朎偺愒摴柺偺嵶朎枌捈壓偵傾僋僠儞僼傿儔儊儞僩偑廤傑傝丄嵶朎枌傪撪懁傊堷偒崬傓偙偲偵傛傝暘楐峚偑偱偒傞偺偼桳柤偱偡丅丂偟偐偟丄傾僋僠儞僼傿儔儊儞僩偲嵶朎枌偑偳偺傛偆側暘巕婡峔偱寢崌偟偰偄傞偐偼丄撲偺傑傑偱偟偨丅丂変乆偑摼偨暘夋偵偼傾僪僿儗儞僗僕儍儞僋僔儑儞偑擹弅偟偰偄傑偟偨丅丂偲偄偆偙偲偼丄傾僋僠儞僼傿儔儊儞僩偲嵶朎枌傪寢崌偝偣傞抈敀幙傕偙偺暘夋偵擹弅偟偰偄傞敜偱偡丅丂偦偺岓曗偲偟偰暘巕検俉俀倠倓偺傾僋僠儞寢崌抈敀幙傪偙偺暘夋偐傜惛惢偟丄儔僨傿僉僔儞乮Radixin乯偲柤晅偗傑偟偨丅丂們俢俶俙僋儘乕僯儞僌傪偟偨偲偙傠丄僄僘儕儞乮Ezrin乯偲屇偽傟傞彫挵偺旝鉕栄偐傜摨掕偝傟偨棤懪偪抈敀幙偲丄儌僄僔儞乮Moesin乯偲屇偽傟傞偁傑傝惈幙偺傛偔暘偐偭偰偄側偐偭偨抈敀幙偲偒傢傔偰傛偔帡偰偄傞偙偲偑暘偐傝傑偟偨丅丂偦偙偱丄塃忋恾偵帵偡傛偆偵丄偦傟偧傟偺摢暥帤傪偲偭偰丄偙偺堚揱巕孮偵ERM僼傽儈儕乕偲柤晅偗傑偟偨丅丂
 傑偨丄偙偺ERM僼傽儈儕乕偼丄僶儞僪係丏侾僗乕僷乕僼傽儈儕亅偵娷傑傟傞偙偲傕暘偐傝傑偟偨丅
傑偨丄偙偺ERM僼傽儈儕乕偼丄僶儞僪係丏侾僗乕僷乕僼傽儈儕亅偵娷傑傟傞偙偲傕暘偐傝傑偟偨丅丂偝偰丄偙偺ERM抈敀幙偼丄攟梴嵶朎偱偼丄俁偮偲傕嫟敪尰偟偰偄偰丄旝鉕栄丄儔僢僼儕儞僌枌丄嵶朎娫偲嵶朎婎幙娫愙拝晹埵丄偝傜偵偼嵍恾偵帵偡傛偆偵暘楐嵶朎偺暘楐峚偵擹弅偟偰偄傑偡丅偙偺偙偲偼丄ERM抈敀幙偑嵶朎枌偲傾僋僠儞僼傿儔儊儞僩偺寢崌偵堦斒揑偵廳梫側栶妱傪偟偰偄傞偙偲傪嫮偔帵嵈偟偰偄傑偟偨丅丂偦傟偧傟偺敪尰傪傾儞僠僙儞僗僆儕僑偱梷偊偰傗傞偲丄旝鉕栄偑側偔側偭偨傝丄愙拝偑夡傟偨傝偟傑偟偨丅丂傑偨丄偙傟傜偺暘巕偼傾儈僲枛抂偱俠俢係係偲屇偽傟傞枌抈敀幙偵捈愙寢崌偡傞偙偲傪尒偮偗傑偟偨丅丂懠偺僌儖乕僾偑僇儖儃僉僔儖枛抂偱傾僋僠儞僼傿儔儊儞僩偵寢崌偡傞偙偲傪帵偟偰偄傑偟偨偺偱丄ERM抈敀幙偑嵶朎枌偲傾僋僠儞僼傿儔儊儞僩偺壦嫶抈敀幙偲偟偰摥偔偙偲偑柧傜偐偵側傝傑偟偨丅
丂偙偺偁偨傝傑偱偼丄ERM抈敀幙偺尋媶偼丄変乆傕娷傔悢僌儖乕僾偑恑傔偰偄傞惷偐側暘栰偱偟偨偑丄堚揱惈偵僔儏儚儞嵶朎偺庮釃偑懡敪偡傞昦婥偱偁傞恄宱慄堐庮徢俀宆乮俶俥俀乯偺偑傫梷惂堚揱巕偑摨掕偝傟丄偦偺嶻暔偑ERM抈敀幙偲偒傢傔偰傛偔帡偰偄傞偙偲偑暘偐傝乮Merlin: Moesin-Ezrin-Radixin-like protein乯丄堦桇媟岝傪梺傃傞寢壥偲側傝傑偟偨丅丂
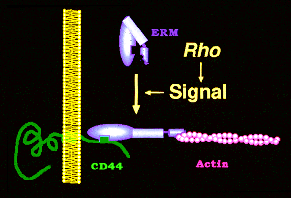 偝傜偵丄塃壓恾偵帵偡傛偆偵丄変乆偑嵶朎枌偲ERM抈敀幙偺寢崌偑掅暘巕検俧俿俹寢崌抈敀幙俼倛倧偵傛偭偰惂屼偝傟偰偄傞偙偲傪戝嶃戝妛偺崅堜尋媶幒偲偲傕偵帵偟丄偦傟偑俼倛倧傪尋媶偟偰偄偨懠偺僌儖乕僾偵傛偭偰傕妋擣偝傟傞帠懺偵帄偭偰丄ERM抈敀幙傪尋媶偡傞僌儖乕僾偼媫懍偵憹偊偮偮偁傝傑偡丅丂偝傜偵丄変乆偼丄ERM抈敀幙偺壦嫶抈敀幙偲偟偰偺妶惈壔忬懺偑椨巁壔偵傛偭偰惂屼偝傟偰偄傞偙偲傪尒弌偟丄傾僋僠儞僼傿儔儊儞僩偲僔僌僫儖揱払偲偺僀儞僞乕僼僃乕僗偲偟偰偺ERM抈敀幙偺夝愅傪懕偗偰偄傑偡丅
偝傜偵丄塃壓恾偵帵偡傛偆偵丄変乆偑嵶朎枌偲ERM抈敀幙偺寢崌偑掅暘巕検俧俿俹寢崌抈敀幙俼倛倧偵傛偭偰惂屼偝傟偰偄傞偙偲傪戝嶃戝妛偺崅堜尋媶幒偲偲傕偵帵偟丄偦傟偑俼倛倧傪尋媶偟偰偄偨懠偺僌儖乕僾偵傛偭偰傕妋擣偝傟傞帠懺偵帄偭偰丄ERM抈敀幙傪尋媶偡傞僌儖乕僾偼媫懍偵憹偊偮偮偁傝傑偡丅丂偝傜偵丄変乆偼丄ERM抈敀幙偺壦嫶抈敀幙偲偟偰偺妶惈壔忬懺偑椨巁壔偵傛偭偰惂屼偝傟偰偄傞偙偲傪尒弌偟丄傾僋僠儞僼傿儔儊儞僩偲僔僌僫儖揱払偲偺僀儞僞乕僼僃乕僗偲偟偰偺ERM抈敀幙偺夝愅傪懕偗偰偄傑偡丅