巹偨偪偺嵶朎娫愙拝憰抲暘夋偐傜惗傑傟偨丄嵟戝偺惉壥偼丄崱偺偲偙傠丄僆僋儖僨傿儞丄僋儘乕僨傿儞偺敪尒偵巒傑傞乽僞僀僩僕儍儞僋僔儑儞偺暘巕惗暔妛揑夝愅乿偱偡丅偙偺堦楢偺尋媶偵傛傝乽僶儕傾乕偺暘巕惗暔妛乿偲偱傕屇傇傋偒丄怴偟偄暘栰偑奐偗偰偒傑偟偨丅偙偺尋媶偵娭偟偰偼丄俀侽侽侽擭偺乽惗壔妛乿偺憤愢丂乮偙偺儁乕僕乯丄俀侽侽侾擭偺乽Nature Review Mol.Cell Biol.乿偺憤愢乮偙偙傪僋儕僢僋乯丄偦偟偰俀侽侽俁擭偺偁傞島墘夛偺婰榐乮偙偙傪僋儕僢僋乯傪偍撉傒壓偝偄丅慡懱憸偑暘偐偭偰偄偨偩偗傞偲巚偄傑偡丅
![]()
乽惗壔妛乿俀侽侽侽擭俁寧崋丂宖嵹
僞僀僩僕儍儞僋僔儑儞傪峔惉偡傞係夞枌娧捠宆抈敀幙
僆僋儖僨傿儞偲僋儘乕僨傿儞偺敪尒丗
Paracellular Pathway偺怴偟偄惗棟妛傊岦偗偰
寧揷彸堦榊丒屆悾姴晇
僞僀僩僕儍儞僋僔儑儞偼丄忋旂嵶朎僔乕僩傗撪旂嵶朎僔乕僩偵偍偄偰丄嵶朎娫傪僔乕儖偡傞廳梫側嵶朎娫愙拝憰抲偱偁傞偑丄堦曽偱丄偙偺僕儍儞僋僔儑儞傪墶愗偭偰暔幙偑嵶朎娫偺捠楬乮paracellular pathway乯傪塣偽傟傞偙偲傕抦傜傟偰偄傞丅丂僞僀僩僕儍儞僋僔儑儞偵偍偄偰椬崌偆嵶朎枌傪僛儘偺嫍棧偵傑偱嬤偯偗偰偄傞愙拝暘巕偺幚懱偼挿偄娫撲偵曪傑傟偰偄偨偑丄嵟嬤偵側偭偰傛偆傗偔丄係夞枌娧捠宆抈敀幙偱偁傞僆僋儖僨傿儞偲僋儘乕僨傿儞偑摨掕偝傟偨丅丂偙偺敪尒偵傛偭偰丄僞僀僩僕儍儞僋僔儑儞偺婡擻傪暘巕惗暔妛揑庤朄傪梡偄偰夝愅偡傞偙偲偑壜擻偲側傝丄怴偟偄paracellular pathway偺惗棟妛偑惗傑傟傛偆偲偟偰偄傞丅
侾丏偼偠傔偵
丂懡悢偺嵶朎偑婡夿揑偵寢傃偮偗傜傟偰偄傞懡嵶朎惗暔偺宍嶌傝偵偍偄偰丄嵶朎娫偍傛傃嵶朎婎幙娫偺愙拝偺壥偨偡栶妱偼戝偒偄丅丂懡條側愙拝暘巕偑丄婡夿揑側寢崌乮愙拝乯偩偗偱側偔丄摿堎揑側嵶朎擣幆傗暋嶨側愙拝僔僌僫儖偺揱払偵傕偐偐傢偭偰偄傞偙偲偑丄師乆偲柧傜偐偵偝傟偰偄傞丅丂偟偐偟丄偙傟傜偲偼慡偔暿偺娤揰偐傜丄嵶朎娫偺愙拝偑懡嵶朎惗暔偵偲偭偰杮幙揑偵廳梫偱偁傞偙偲偑屆偔偐傜抦傜傟偰偄傞丅丂偦傟偼丄乽嵶朎娫傪捠偭偨暔幙偺堏摦乮楻傟乯偺惂屼乿偲偄偆娤揰偱偁傞丅
丂懡嵶朎惗暔偼忋旂嵶朎偵埻傑傟傞偙偲偵傛傝丄傑偢丄帺屓偺撪偲奜偵嬫暿偝傟傞丅丂偦偟偰丄懱偺拞偼丄忋旂嵶朎傗撪旂嵶朎偺僔乕僩偵傛傝丄偝傜偵偄偔偮傕偺晹壆偵暘偗傜傟偰偄傞丅丂擼丄寣娗丄恡憻偺擜嵶娗側偳偼偦偺戙昞揑側晹壆偱偁傞偑丄摿庩側傕偺偱偼峛忬態偺鄅朎丄撪帹偺鍡媿娗側偳傕嫇偘傜傟傞丅丂晹壆偺拞偺僀僆儞娐嫬傗抈敀幙偺庬椶丒擹搙側偳偼丄偦傟偧傟偺婡擻偵墳偠偰戝偒偔堎側偭偰偍傝丄偙偺娐嫬傪摦揑偵曐偭偰偄傞偙偲偑丄懡嵶朎惗暔偑惗偒偰偄偔忋偱昁梫晄壜寚偱偁傞丅丂偟偐偟丄懡嵶朎惗暔偱偁傞偑屘偵丄偙傟傜偺晹壆傪巇愗傞暻偼嵶朎傪暲傋偰嶌傜偞傞傪摼側偄偑丄偄偔傜僇僪僿儕儞側偳偺嫮椡側愙拝暘巕偱嵶朎娫傪寢崌偝偣偰傕丄悈丒僀僆儞丒抈敀幙側偳偼嵶朎娫傪帺桼偵捠傞偙偲偑偱偒傞丅丂偦偙偱丄懡嵶朎惗暔偑懚嵼偡傞偨傔偵偼丄偙偺嵶朎娫傪捠偭偨暔幙偺堏摦乮楻傟乯傪杊偖偨傔偺摿庩側愙拝婡峔偑昁梫偲側傞丅丂丂偦傟偑屻弎偡傞僞僀僩僕儍儞僋僔儑儞乮埲壓俿俰乯偲屇偽傟傞愙拝憰抲偱丄偙偙偱偼椬傝崌偆嵶朎偺嵶朎枌偺嫍棧偑僛儘偵傑偱嬤偯偄偰偄偰丄嵶朎娫傪捠偭偨暔幙偺堏摦傪杊偄偱偄傞丅
丂愐捙摦暔偱偼丄偙偺俿俰傪忋旂嵶朎娫傗撪旂嵶朎娫偵敪払偝偣傞偙偲偵傛傝丄偙傟傜偺嵶朎僔乕僩偑奺晹壆偺娐嫬傪庣傞僶儕傾乕偲偟偰摥偔偙偲傪壜擻偲偟偰偄傞丅丂偟偐偟丄愭偵傕弎傋偨傛偆偵丄奺晹壆偺撪晹偺娐嫬偼惷揑偵曐偨傟偰偄傞偺偱偼側偔丄晹壆偺奜偲暔幙偺岎姺傪偟側偑傜摦揑偵曐偨傟偰偄傞丅丂偙偺嵺丄偄傠偄傠側暔幙偑嵶朎僔乕僩傪墶愗傞昁梫偑偁傞偑丄偦傟偵偼恾侾偵帵偡傛偆側俀偮偺儖乕僩偑偁傞丅丂嵶朎偦偺傕偺丄偡側傢偪嵶朎枌偦偺傕偺傪墶愗偭偰塣偽傟傞儖乕僩偼transcellular
pathway偲屇偽傟丄偙偙偵偼懡偔偺僠儍僱儖暘巕傗億儞僾丄僩儔儞僗億乕僞乕側偳偑偐偐傢偭偰偄傞丅
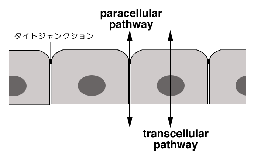 丂偙偺pathway偼偙傟傑偱偺惗棟妛尋媶偺拞怱揑僥乕儅偱偁偭偨丅丂堦曽丄嵶朎娫丄偡側傢偪俿俰傪墶愗偭偰塣偽傟傞儖乕僩偼paracellualr
pathway偲屇偽傟傞丅丂俿俰偑僶儕傾乕偺偨傔偵懚嵼偡傞偲偄偆丄愭偺媍榑偲堦尒柕弬偡傞傛偆偱偁傞偑丄幚嵺偵偼俿俰偺僶儕傾乕婡擻偼丄偒傢傔偰tight側傕偺偐傜丄偐側傝leaky側傕偺傑偱憡摉偺暆偑偁傝丄堦斒揑偵俿俰偼慖戰揑偵僀僆儞側偳傪捠偟偆傞僶儕傾乕偱偁傞偲尵偆偙偲偑偱偒傞丅丂丂偡側傢偪俿俰偼paracellular
channel偲偱傕尵偆傋偒乬寠乭傪撪曪偡傞偲峫偊傜傟傞偺偱偁傞1)丅丂暔幙偵傛偭偰偼庡偵paracellular pathway傪捠偭偰晹壆偺撪奜偱岎姺偝傟傞偙偲傕抦傜傟偰偍傝丄偙偺pathway傕transcellualr
pathway偲摨條偵廳梫偱偁傞偲峫偊傜傟偰偒偨丅丂偟偐偟丄俿俰偺暘巕揑婎斦偵娭偡傞変乆偺抦幆偑偒傢傔偰枹弉偱偁偭偨偨傔偵丄偦偺夝愅偼抶傟偰偄偨2)丅
丂偙偺pathway偼偙傟傑偱偺惗棟妛尋媶偺拞怱揑僥乕儅偱偁偭偨丅丂堦曽丄嵶朎娫丄偡側傢偪俿俰傪墶愗偭偰塣偽傟傞儖乕僩偼paracellualr
pathway偲屇偽傟傞丅丂俿俰偑僶儕傾乕偺偨傔偵懚嵼偡傞偲偄偆丄愭偺媍榑偲堦尒柕弬偡傞傛偆偱偁傞偑丄幚嵺偵偼俿俰偺僶儕傾乕婡擻偼丄偒傢傔偰tight側傕偺偐傜丄偐側傝leaky側傕偺傑偱憡摉偺暆偑偁傝丄堦斒揑偵俿俰偼慖戰揑偵僀僆儞側偳傪捠偟偆傞僶儕傾乕偱偁傞偲尵偆偙偲偑偱偒傞丅丂丂偡側傢偪俿俰偼paracellular
channel偲偱傕尵偆傋偒乬寠乭傪撪曪偡傞偲峫偊傜傟傞偺偱偁傞1)丅丂暔幙偵傛偭偰偼庡偵paracellular pathway傪捠偭偰晹壆偺撪奜偱岎姺偝傟傞偙偲傕抦傜傟偰偍傝丄偙偺pathway傕transcellualr
pathway偲摨條偵廳梫偱偁傞偲峫偊傜傟偰偒偨丅丂偟偐偟丄俿俰偺暘巕揑婎斦偵娭偡傞変乆偺抦幆偑偒傢傔偰枹弉偱偁偭偨偨傔偵丄偦偺夝愅偼抶傟偰偄偨2)丅
2. 僞僀僩僕儍儞僋僔儑儞偺峔憿
丂 扨憌偺忋旂嵶朎偱偼丄椬傝崌偆嵶朎枌乮儔僥儔儖枌乯偺傕偭偲傕傾僺僇儖傛傝偵嵶朎娫愙拝憰抲暋崌懱偲屇偽傟傞椞堟偑偁傝丄偦偺晹暘偱椬傝崌偆嵶朎偑嫮偔愙拝偟偰偄傞乮恾俀俙乯丅丂偙偺椞堟偼俁偮偵暘偐傟丄傾僺僇儖懁偐傜僞僀僩僕儍儞僋僔儑儞乮俿俰乯丄傾僪僿儗儞僗僕儍儞僋僔儑儞乮俙俰乯丄僨僗儌僜乕儉乮俢俽乯偲屇偽傟傞3)丅丂俙俰偵偼僇僪僿儕儞偑愙拝暘巕偲偟偰擹弅偟偰偍傝丄傛偔敪払偟偨棤懪偪峔憿傪夘偟偰傾僋僠儞僼傿儔儊儞僩偑枾偵嵶朎枌偵寢崌偟偰偄傞丅丂俢俽偱傕傗偼傝僇僪僿儕儞條偺愙拝暘巕偑婡擻偟偰偍傝丄偙偙
 偱偼拞娫宎僼傿儔儊儞僩偑寢崌偟偰偄傞丅丂偙傟傜俀偮偺愙拝憰抲偱偼岦偐偄崌偆嵶朎枌娫偺嫍棧偼15-20 nm偵曐偨傟偰偄傞丅丂嵶朎娫偺嫮偄婡夿揑寢崌傪扴偭偰偍傝丄偦偺寢崌椡傪嵶朎崪奿偵偮側偘傞偙偲偵傛傝丄慻怐慡懱偲偟偰偺婡夿揑嫮搙傪傕扴偭偰偄傞丅丂偙傟傜偵妑傋丄俿俰偼摿庩側愙拝憰抲偱偁傞丅丂挻敄愗曅憸偱尒傞偲丄偲偙傠偳偙傠偱岦偐偄崌偆嵶朎枌娫偺嫍棧偑僛儘偵傑偱嬤偯偄偰偄傞3)乮俿俰偺僉僢僔儞僌億僀儞僩偲屇偽傟傞乯乮恾俀俛乯丅丂傑偨丄搥寢妱抐儗僾儕僇朄偱娤嶡偡傞偲丄俿俰偺晹暘偱偼丄枌撪棻巕偑堦楍偵暲傫偩僗僩儔儞僪乮俿俰僗僩儔儞僪乯偑曇傒栚傪峔惉偟偰偄傞4)乮恾俀俠乯丅丂偙偺傛偆側娤嶡偐傜丄俿俰偺峔憿偼恾俀俙偺傛偆側傕偺偱偁傞偲憐憸偝傟偰偒偨丅丂偡側傢偪丄壗傜偐偺撪嵼惈枌抈敀幙偑嵶朎枌偺拞偱慄忬偵廳崌偟偰俿俰僗僩儔儞僪傪宍惉偟丄岦偐偄崌偆嵶朎枌拞偺俿俰僗僩儔儞僪偲懁柺偱懳崌偡傞偙偲偵傛傝僉僢僔儞僌億僀儞僩傪宍惉偡傞偲偄偆峔憿偱偁傞丅丂偟偐偟丄偙偺俿俰僗僩儔儞僪傪峔惉偡傞撪嵼惈枌抈敀幙偺幚懱偼挿偄娫撲偵曪傑傟偰偍傝丄俿俰僗僩儔儞僪偑摿庩側帀幙偺媡儈僙儖偐傜弌棃偰偄傞偲偄偆帀幙愢傕桳椡帇偝傟偰懕偗偰偒偨5)丅
偱偼拞娫宎僼傿儔儊儞僩偑寢崌偟偰偄傞丅丂偙傟傜俀偮偺愙拝憰抲偱偼岦偐偄崌偆嵶朎枌娫偺嫍棧偼15-20 nm偵曐偨傟偰偄傞丅丂嵶朎娫偺嫮偄婡夿揑寢崌傪扴偭偰偍傝丄偦偺寢崌椡傪嵶朎崪奿偵偮側偘傞偙偲偵傛傝丄慻怐慡懱偲偟偰偺婡夿揑嫮搙傪傕扴偭偰偄傞丅丂偙傟傜偵妑傋丄俿俰偼摿庩側愙拝憰抲偱偁傞丅丂挻敄愗曅憸偱尒傞偲丄偲偙傠偳偙傠偱岦偐偄崌偆嵶朎枌娫偺嫍棧偑僛儘偵傑偱嬤偯偄偰偄傞3)乮俿俰偺僉僢僔儞僌億僀儞僩偲屇偽傟傞乯乮恾俀俛乯丅丂傑偨丄搥寢妱抐儗僾儕僇朄偱娤嶡偡傞偲丄俿俰偺晹暘偱偼丄枌撪棻巕偑堦楍偵暲傫偩僗僩儔儞僪乮俿俰僗僩儔儞僪乯偑曇傒栚傪峔惉偟偰偄傞4)乮恾俀俠乯丅丂偙偺傛偆側娤嶡偐傜丄俿俰偺峔憿偼恾俀俙偺傛偆側傕偺偱偁傞偲憐憸偝傟偰偒偨丅丂偡側傢偪丄壗傜偐偺撪嵼惈枌抈敀幙偑嵶朎枌偺拞偱慄忬偵廳崌偟偰俿俰僗僩儔儞僪傪宍惉偟丄岦偐偄崌偆嵶朎枌拞偺俿俰僗僩儔儞僪偲懁柺偱懳崌偡傞偙偲偵傛傝僉僢僔儞僌億僀儞僩傪宍惉偡傞偲偄偆峔憿偱偁傞丅丂偟偐偟丄偙偺俿俰僗僩儔儞僪傪峔惉偡傞撪嵼惈枌抈敀幙偺幚懱偼挿偄娫撲偵曪傑傟偰偍傝丄俿俰僗僩儔儞僪偑摿庩側帀幙偺媡儈僙儖偐傜弌棃偰偄傞偲偄偆帀幙愢傕桳椡帇偝傟偰懕偗偰偒偨5)丅
俁丏僆僋儖僨傿儞偺摨掕偲婡擻夝愅
丂変乆偼丄傕偲傕偲丄僇僪僿儕儞偑婡擻偡傞俙俰偺暘巕峔抸傪柧傜偐偵偡傞偨傔偵丄俙俰偑擹弅偟偨暘夋傪娞憻偐傜扨棧偡傞曽朄傪奐敪偟偰偄偨6)丅丂偦偟偰丄徻嵶偼徣棯偡傞偑丄倅俷亅侾偲屇偽傟傞嵶朎枌棤懪偪抈敀幙偺們俢俶俙僋儘乕僯儞僌偺夁掱偱丄偙偺暘夋偵俿俰傕嬌搙偵擹弅偟偰偄傞偙偲偵婥偯偄偨7)丅丂傕偟丄俿俰僗僩儔儞僪偑撪嵼惈枌抈敀幙偵傛偭偰峔惉偝傟偰偄傞偺偱偁傟偽丄偦偺傛偆側抈敀幙傕偙偺暘夋偵擹弅偟偰偄傞敜偱偁傞丅丂偦偙偱偙偺扨棧暘夋傪峈尨偵偟偰儌僲僋儘乕僫儖峈懱傪摼傞偙偲偵傛傝丄俿俰僗僩儔儞僪傪峔惉偡傞枌抈敀幙偺摨掕傪帋傒傞偙偲偵偟偨丅丂幚嵺偵偼崅偄峈尨惈傪摼傞偨傔偵丄僸儓僐偺娞憻偐傜暘夋傪摼偰丄偦傟傪儔僢僩偵柶塽偡傞偙偲偵傛傝懡悢偺儌僲僋儘乕僫儖峈懱傪摼偨丅丂偦偺拞偐傜俿俰偺枌抈敀幙傪擣幆偡傞壜擻惈偺偁傞儌僲僋儘乕僫儖峈懱傪峣傝崬傫偩偲偙傠丄暘巕検俇俆倠俢倎偖傜偄偺暆峀偄僶儞僪傪擣幆偡傞傕偺偑俁庬椶偲傟偨丅丂偙傟傜偺峈懱偼揹巕尠旝嬀儗儀儖偱俿俰僗僩儔儞僪偦偺傕偺傪擣幆偟偨丅丂丂傑偨丄偦偺峈尨偑撪嵼惈枌抈敀幙偱偁傞偙偲傕惗壔妛揑偵妋擣偱偒偨丅丂丂偦偙偱偙偺峈尨傪僆僋儖僨傿儞乮occludin丗儔僥儞岅偺occlude丄暵偠傞偺堄丄偐傜乯偲柤晅偗丄偦偺們俢俶俙傪扨棧偟偨偲偙傠丄怴婯偺係夞枌娧捠宆抈敀幙偱偁傞偙偲偑柧傜偐偵側偭偨8)丅丂恾俁偵帵偡傛偆偵丄僆僋儖僨傿儞偼偦偺N枛抂懁敿暘偵偦傟偧傟栺係侽傾儈僲巁偐傜側傞俀偮偺嵶朎奜儖乕僾傪帩偮偑丄摿偵戞堦儖乕僾偺栺俈侽亾偺傾儈僲巁偑僠儘僔儞偐僌儕僔儞偲偄偆婏柇側峔憿傪偟偰偄偨丅丂偙偺傛偆偵偟偰丄俿俰僗僩儔儞僪傪峔惉偡傞枌抈敀幙偲偟偰丄僯儚僩儕偺僆僋儖僨傿儞偑弶傔偰摨掕偝傟偨偺偼丄侾俋俋俁擭偺曢傟偺偙偲偱丄俿俰偺峔憿偲婡擻偺棟夝偑堦婥偵恑傓偐偲巚傢傟偨2)丅丂偟偐偟丄幚嵺偵偼丄僆僋儖僨傿儞偺傾儈僲巁攝楍偑庬娫偱戝偒偔曄摦偟偰偄偨偨傔偵丄儅僂僗傗僸僩偺僆僋儖僨傿儞偺摨掕偼擄峲偟偨丅丂彊乆偵廩幚偟偰偒偨堚揱巕偺僨乕僞儀乕僗偺拞偐傜丄変乆偑岾塣偵傕僸僩偺僆僋儖僨傿儞偺堦晹偺攝楍傪尒偮偗丄傛偆傗偔傎擕椶偺僆僋儖僨傿儞偺摨掕偵惉岟偡傞傑偱偵栺俀擭偺帪娫偑昁梫偱偁偭偨9)丅
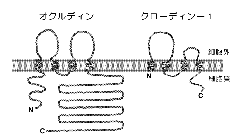
丂堦曽偱丄僯儚僩儕偱偼偁傞偑丄俿俰偺撪嵼惈枌抈敀幙偑尒偮偐偭偨偲偄偆偙偲偱丄懡偔偺尋媶幒偱僆僋儖僨傿儞偺婡擻夝愅偑巒傑偭偨丅丂偦偺寢壥丄僆僋儖僨傿儞偑俿俰僗僩儔儞僪偺宍惉傪捠偟偰丄俿俰偺婡擻傪捈愙扴偭偰偄傞偲偄偆偙偲傪庡挘偡傞榑暥偑師乆偲敪昞偝傟傞傛偆偵側偭偨10)丅丂偟偐偟丄変乆偼丄杮摉偵僆僋儖僨傿儞偩偗偱俿俰僗僩儔儞僪偑宍惉偝傟偰偄傞偺偐偳偆偐偵偮偄偰丄彊乆偵媈栤傪帩偮傛偆偵側偭偰偒偨丅丂偙偺傛偆側媈栤偵捈愙摎偊傞堦偮偺曽朄偼丄僆僋儖僨傿儞偺堚揱巕傪僲僢僋傾僂僩偡傞偙偲偱偁傞丅丂偦偙偱丄儅僂僗偺僆僋儖僨傿儞堚揱巕偺扨棧偵惉岟偟偨屻偵丄憡摨慻傒姺偊傪棙梡偟偰俤俽乮embryonic
stem乯嵶朎偵偍偄偰僆僋儖僨傿儞偺堚揱巕傪僟僽儖僲僢僋傾僂僩偟偨丅丂偦偺嵶朎傪帋尡娰撪偱忋旂嵶朎偵暘壔偝偣偨偲偙傠丄梊憐奜側偙偲偵丄僆僋儖僨傿儞傪敪尰偟側偄偙偺傛偆側忋旂嵶朎偱傕棫攈側俿俰僗僩儔儞僪偺僱僢僩儚乕僋偑宍惉偝傟偨11)丅丂偙偺帠幚偼丄僆僋儖僨傿儞偑側偔偰傕俿俰僗僩儔儞僪偑宍惉偝傟傞偲偄偆偙偲傪堄枴偟偰偄偨丅
係丏僋儘乕僨傿儞亅侾丄亅俀偺摨掕
丂偙偺傛偆偵堚揱巕傪攋夡偟偰傕僼僃僲僞僀僾偵曄壔偑側偄応崌偼丄僆僋儖僨傿儞堚揱巕偑僼傽儈儕乕傪宍惉偟偰偄偰丄懠偺儊儞僶乕偑戙傢傝傪偡傞偨傔偲峫偊傞偺偑晛捠偱偁傞丅丂偟偐偟丄僆僋儖僨傿儞偵娭偟偰偼丄僨乕僞儀乕僗忋傕丄俼俿亅俹俠俼摍偵傛傞専嶕忋傕丄帡偨傾儈僲巁攝楍傪帩偮枌抈敀幙偼尒偮偐傜側偐偭偨丅丂偦偙偱丄僆僋儖僨傿儞偲壗傜偐偺堄枴偱憡屳嶌梡偡傞枌抈敀幙傪扵偡偲偄偆曽岦偱丄偙偺僷儔僪僢僋僗傪夝偙偆偲峫偊偨丅丂yeast
two-hybrid朄傗柶塽捑崀朄傪帋傒偨偑惉岟偣偢丄忋婰偺僕儍儞僋僔儑儞偺扨棧暘夋偵擹弅偡傞枌抈敀幙偺拞偱丄庬乆偺忦審壓乮挻壒攇攋夡傗潋摐枾搙岡攝墦怱乯偱僆僋儖僨傿儞偲嫇摦傪偲傕偵偡傞傕偺傪扵偡偙偲偵偟偨丅丂偦偺寢壥丄嵟廔揑偵丄揹婥塲摦忋俀俁倠俢倎偺暘巕検傪帵偡抈敀幙偑岓曗偲偟偰巆偭偨偺偱丄偦偺傾儈僲巁攝楍傪寛掕偟偨偲偙傠丄俀偮偺俤俽俿僋儘乕儞偵堦抳偟偨丅丂偙傟傜偺俤俽俿僋儘乕儞偐傜慡挿偺堚揱巕傪扨棧偟偨偲偙傠丄屳偄偵傾儈僲巁儗儀儖偱栺俁侽亾偺儂儌儘僕乕傪帵偡俀庬椶偺俀俁倠俢倎抈敀幙傪僐乕僪偡傞們俢俶俙偑摨掕偝傟偨乮傕偲傕偲偺俀俁倠俢倎偺僶儞僪偑彮側偔偲傕俀庬椶埲忋偺抈敀幙偺崿崌暔偩偭偨偙偲偵側傞乯丅丂偍傕偟傠偄偙偲偵丄偙傟傜偺抈敀幙偼偦偺傾儈僲巁攝楍偐傜丄傗偼傝係夞枌娧捠宆偺撪嵼惈枌抈敀幙偲巚傢傟偨偑丄僆僋儖僨傿儞偲偼慡偔儂儌儘僕乕傪帵偝側偐偭偨
乮恾俁乯丅丂  偄偢傟偺抈敀幙傕僞僌傪偮偗偰忋旂嵶朎偵敪尰偝偣傞偲俿俰僗僩儔儞僪偵庢傝崬傑傟傞偙偲偑暘偐偭偨偺偱丄偙偺帪揰偱僋儘乕僨傿儞亅侾偲亅俀乮claudin-1,-2丗儔僥儞岅偺claudere丄暵偠傞偺堄丄偐傜乯偲柤晅偗偨12)丅
偄偢傟偺抈敀幙傕僞僌傪偮偗偰忋旂嵶朎偵敪尰偝偣傞偲俿俰僗僩儔儞僪偵庢傝崬傑傟傞偙偲偑暘偐偭偨偺偱丄偙偺帪揰偱僋儘乕僨傿儞亅侾偲亅俀乮claudin-1,-2丗儔僥儞岅偺claudere丄暵偠傞偺堄丄偐傜乯偲柤晅偗偨12)丅
丂僋儘乕僨傿儞亅侾傑偨偼亅俀傪俿俰傪帩偨側偄儅僂僗俴慄堐夎嵶朎偵嫮惂敪尰偝偣傞偲丄偄偢傟偺応崌傕丄傕偲傕偲嵶朎娫愙拝妶惈傪帵偝側偄俴嵶朎偑屳偄偵嫮屌偵愙拝偡傞傛偆偵側偭偨13)丅丂偝傜偵嵶朎娫偵嫄戝側俿俰僗僩儔儞僪僱僢僩儚乕僋偑宍惉偝傟偨乮恾係乯丅丂僆僋儖僨傿儞傪俴嵶朎偵敪尰偝偣偰傕偙偺傛偆側僱僢僩儚乕僋偼宍惉偝傟側偄偑丄僋儘乕僨傿儞偲摨帪偵摫擖偡傞偲丄僋儘乕僨傿儞偵傛偭偰宍惉偝傟偨僗僩儔儞僪偵僆僋儖僨傿儞偑慻傒崬傑傟偨14)丅
俆丏僋儘乕僨傿儞堚揱巕僼傽儈儕乕
丂僋儘乕僨傿儞亅侾偲亅俀偺懚嵼偼丄僋儘乕僨傿儞堚揱偑僼傽儈儕乕傪宍惉偟偰偄傞偙偲傪堄枴偟偰偄偨丅丂偦偙偱僨乕僞儀乕僗傪挷傋偰傒傞偲丄僋儘乕僨傿儞亅侾偲亅俀偵帡偨傾儈僲巁攝楍偑戲嶳搊榐偝傟偰偄偨丅丂
偙傟傜偺攝楍傪傕偲偵丄偦傟偧傟傪僐乕僪偡傞堚揱巕傪摨掕偟偰偄偭偨偲偙傠丄嵟廔揑偵僋儘乕僨傿儞僼傽儈儕乕偼儅僂僗偱彮側偔偲傕侾俆庬椶偺儊儞僶乕偐傜側傞偙偲偑柧傜偐偵側偭偨15,16)丅丂
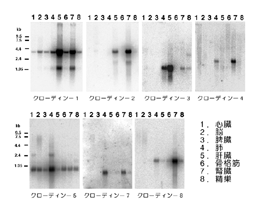 恾俆偼丄僋儘乕僨傿儞亅侾偐傜亅俉傑偱偺奺憻婍偵偍偗傞敪尰傪僲僓儞僽儘僢僩偵傛傝挷傋偨傕偺偱偁傞偑丄偦傟偧傟偺僋儘乕僨傿儞偑暋嶨側慻傒崌傢偣偱奺憻婍偱敪尰偟偰偄傞偺偑暘偐傞丅丂椺偊偽丄僋儘乕僨傿儞亅侾偼斾妑揑懡偔偺憻婍偱敪尰偟偰偄傞偑丄摿偵娞憻偲恡憻偵懡偄丅丂堦曽丄僋儘乕僨傿儞亅俁偺敪尰偼攛偲娞憻偵尷傜傟偰偄傞丅丂傑偨丄僋儘乕僨傿儞亅俇偼惉懱偱偼敪尰偑尒傜傟偢丄戀帣偱偼懡偔偺憻婍偵敪尰偟偰偄傞傛偆偱偁傞丅
恾俆偼丄僋儘乕僨傿儞亅侾偐傜亅俉傑偱偺奺憻婍偵偍偗傞敪尰傪僲僓儞僽儘僢僩偵傛傝挷傋偨傕偺偱偁傞偑丄偦傟偧傟偺僋儘乕僨傿儞偑暋嶨側慻傒崌傢偣偱奺憻婍偱敪尰偟偰偄傞偺偑暘偐傞丅丂椺偊偽丄僋儘乕僨傿儞亅侾偼斾妑揑懡偔偺憻婍偱敪尰偟偰偄傞偑丄摿偵娞憻偲恡憻偵懡偄丅丂堦曽丄僋儘乕僨傿儞亅俁偺敪尰偼攛偲娞憻偵尷傜傟偰偄傞丅丂傑偨丄僋儘乕僨傿儞亅俇偼惉懱偱偼敪尰偑尒傜傟偢丄戀帣偱偼懡偔偺憻婍偵敪尰偟偰偄傞傛偆偱偁傞丅
丂偄偔偮偐偺摿庩側嵶朎偼丄摿庩側僋儘乕僨傿儞傪敪尰偟偰偄傞偲偄偆偙偲傕丄嵟嬤柧傜偐偵側偭偰偒偨丅丂椺偊偽丄恾俆偵尒傜傟傞傛偆偵僋儘乕僨傿儞亅俆偼丄擼丄怱嬝丄崪奿嬝偲偄偭偨忋旂嵶朎傪娷傑側偄憻婍偵傕懡偔敪尰偟偰偄傞丅丂偙偺偙偲偐傜丄偙偺僋儘乕僨傿儞偼寣娗偺撪旂嵶朎偵敪尰偟偰偄傞偺偱偼側偄偐偲峫偊丄摿堎峈懱偺嶌惢傪帋傒偨丅偦偺寢壥丄梊憐捠傝丄僋儘乕僨傿儞亅俆偑寣娗摿堎揑僇僪僿儕儞偱偁傞倁俤僇僪僿儕儞偲偲傕偵丄寣娗偺撪旂嵶朎娫偵擹弅偟偰偄傞偙偲偑柧傜偐偵側偭偨17)乮恾俇乯丅丂
崱偺偲偙傠丄懠偺庬椶偺僋儘乕僨傿儞偺敪尰偼寣娗撪旂嵶朎偵偼尒傜傟偰偄側偄偺偱丄 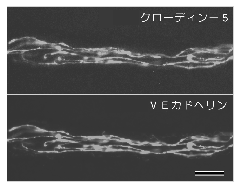 僋儘乕僨傿儞亅俆偑寣娗偺摟夁惈傪婯掕偡傞廳梫側暘巕偱偁傞壜擻惈偑崅偄丅丂傕偆堦偮偺椺偲偟偰丄僋儘乕僨傿儞乕侾侾偑嫇偘傜傟傞丅丂偙偺僋儘乕僨傿儞偼丄僲僓儞僽儘僢僩偱偼庡偵擼偲惛憙偵敪尰偟偰偄傞丅丂傗偼傝摿堎峈懱傪嶌惢偟偰挷傋偨偲偙傠丄擼偱偼儈僄儕儞偺嵶朎枌娫偺摿庩側俿俰傪丄惛憙偱偼僙儖僩儕嵶朎娫偺傛偔敪払偟偨俿俰傪峔惉偟偰偄傞偙偲偑暘偐偭偨18)丅丂偡側傢偪丄偙偺僋儘乕僨傿儞偼丄桳悜恄宱偵偍偗傞挼桇揱摫傗丄惛憙偵偍偗傞寣娗惛憙僶儕傾乕偵偲偭偰廳梫側暘巕偱偁傞偙偲偑帵嵈偝傟偨丅
僋儘乕僨傿儞亅俆偑寣娗偺摟夁惈傪婯掕偡傞廳梫側暘巕偱偁傞壜擻惈偑崅偄丅丂傕偆堦偮偺椺偲偟偰丄僋儘乕僨傿儞乕侾侾偑嫇偘傜傟傞丅丂偙偺僋儘乕僨傿儞偼丄僲僓儞僽儘僢僩偱偼庡偵擼偲惛憙偵敪尰偟偰偄傞丅丂傗偼傝摿堎峈懱傪嶌惢偟偰挷傋偨偲偙傠丄擼偱偼儈僄儕儞偺嵶朎枌娫偺摿庩側俿俰傪丄惛憙偱偼僙儖僩儕嵶朎娫偺傛偔敪払偟偨俿俰傪峔惉偟偰偄傞偙偲偑暘偐偭偨18)丅丂偡側傢偪丄偙偺僋儘乕僨傿儞偼丄桳悜恄宱偵偍偗傞挼桇揱摫傗丄惛憙偵偍偗傞寣娗惛憙僶儕傾乕偵偲偭偰廳梫側暘巕偱偁傞偙偲偑帵嵈偝傟偨丅
俇丏僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪偺暘巕峔抸
丂慜弎偟偨傛偆偵丄俀杮偺俿俰僗僩儔儞僪偑懁柺偱懳崌偡傞偙偲偵傛傝俿俰偺僉僢僔儞僌億僀儞僩偑宍惉偝傟偰偄傞丅丂
堦曽丄俴嵶朎偵僩儔儞僗僼僃僋僔儑儞偡傞幚尡偵傛傝丄僋儘乕僨傿儞偑慄忬偵廳崌偟偰俿俰僗僩儔儞僪偑宍惉偝傟傞偙偲偑徹柧偝傟偰偄傞13)丅丂暋悢偺庬椶偺僋儘乕僨傿儞偑堦偮偺嵶朎偵嫟敪尰偟偰偄傞偲側傞偲丄懳崌偟偰偄傞俀杮偺僗僩儔儞僪偺拞偵偙傟傜偺僋儘乕僨傿儞偼偳偺傛偆偵慻傒崬傑傟偰偄傞偺偱偁傠偆偐丠丂栤戣偑暋嶨偵側傞偺偱丄恾俈偱偼俀庬椶偺僋儘乕僨傿儞偑嫟敪尰偟偰偄傞応崌偵偮偄偰峫偊偰傒傞丅丂
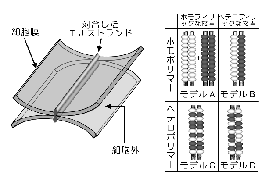 傑偢丄偦傟偧傟偺僗僩儔儞僪偑摨堦偺僋儘乕僨傿儞偐傜側傞儂儌億儕儅乕偱偁傞働乕僗偲丄俀庬椶偺僋儘乕僨傿儞偑嫟廳崌偟偰乮僿僥儘億儕儅乕乯堦杮偺僗僩儔儞僪傪宍惉偟偰偄傞働乕僗偑峫偊傜傟傞丅丂偝傜偵丄岦偐偄崌偆僗僩儔儞僪偳偆偟偑懳崌偡傞嵺丄摨偠僋儘乕僨傿儞偳偆偟偑愙拝偡傞働乕僗乮儂儌僼傿儕僢僋側愙拝乯偲丄堎側傞僋儘乕僨傿儞偳偆偟傕愙拝偱偒傞働乕僗乮僿僥儘僼傿儕僢僋側愙拝乯偑峫偊傜傟傞丅丂偙傟傜傪傑偲傔傞偲丄扨弮偵偼俙偐傜俢傑偱偺係庬椶偺儌僨儖偑峫偊傜傟傞丅丂幚嵺偺俿俰偱偼丄偳偺儌僨儖偑摉偰偼傑傞偺偱偁傠偆偐丠
傑偢丄偦傟偧傟偺僗僩儔儞僪偑摨堦偺僋儘乕僨傿儞偐傜側傞儂儌億儕儅乕偱偁傞働乕僗偲丄俀庬椶偺僋儘乕僨傿儞偑嫟廳崌偟偰乮僿僥儘億儕儅乕乯堦杮偺僗僩儔儞僪傪宍惉偟偰偄傞働乕僗偑峫偊傜傟傞丅丂偝傜偵丄岦偐偄崌偆僗僩儔儞僪偳偆偟偑懳崌偡傞嵺丄摨偠僋儘乕僨傿儞偳偆偟偑愙拝偡傞働乕僗乮儂儌僼傿儕僢僋側愙拝乯偲丄堎側傞僋儘乕僨傿儞偳偆偟傕愙拝偱偒傞働乕僗乮僿僥儘僼傿儕僢僋側愙拝乯偑峫偊傜傟傞丅丂偙傟傜傪傑偲傔傞偲丄扨弮偵偼俙偐傜俢傑偱偺係庬椶偺儌僨儖偑峫偊傜傟傞丅丂幚嵺偺俿俰偱偼丄偳偺儌僨儖偑摉偰偼傑傞偺偱偁傠偆偐丠
丂娞憻偺幚幙嵶朎偱偼丄彮側偔偲傕僋儘乕僨傿儞亅侾丄亅俀丄亅俁偺俁庬椶偺僋儘乕僨傿儞偑嫟敪尰偟偰偄傞丅丂揹巕尠旝嬀儗儀儖偺峈懱愼怓偱偼丄偙傟傜偺僋儘乕僨傿儞偑俿俰僗僩儔儞僪偵斾妑揑嬒堦偵崿偞傝崌偭偰偄傞丄偡側傢偪丄僿僥儘億儕儅乕傪宍惉偟偰偄傞傛偆偵傒偊傞丅丂幚嵺丄
俴嵶朎偵偙傟傜偺僋儘乕僨傿儞偺偆偪丄擟堄偺俀庬椶傪嫟敪尰偝偣傞偲丄妋偐偵僿僥儘億儕儅乕傪嶌傞偙偲偑徹柧偝傟偨19)丅丂傕偪傠傫丄僋儘乕僨傿儞亅侾傗亅俀傪扨撈偱敪尰偝偣偰傕俿俰僗僩儔儞僪偑嵞峔惉偝傟傞偺偱丄儂儌億儕儅乕傕嶌傝摼傞13)丅丂偦偙偱丄師偵丄僋儘乕僨傿儞亅侾偩偗丄傑偨偼僋儘乕僨傿儞亅俀偩偗傪敪尰偝偣偨俴嵶朎乮偦傟偧傟俠侾俴嵶朎偍傛傃俠俀俴嵶朎乯傪丄僋儘乕僨傿儞亅俁偩偗傪敪尰偝偣偨俴嵶朎乮俠俁俴嵶朎乯偲崿偤偰攟梴偟偰傒偨偲偙傠丄俠侾俴乛俠俁俴嵶朎娫偵傕俠俀俴乛俠俁俴嵶朎娫偵傕傛偔敪払偟偨俿俰僗僩儔儞僪僱僢僩儚乕僋偑宍惉偝傟偨丅丂偡側傢偪丄僋儘乕僨傿儞亅侾偲亅俁丄僋儘乕僨傿儞亅俀偲亅俁偼丄偦傟偧傟僿僥儘僼傿儕僢僋偵愙拝偡傞偙偲偵傛傝懳崌偟偨俿俰僗僩儔儞僪傪宍惉偡傞偙偲偑偱偒傞偺偱偁傞丅丂偟偨偑偭偰丄係偮偺儌僨儖偺偆偪丄傕偭偲傕暋嶨側儌僨儖俢偑幚嵺偺俿俰偱婲偒偰偄傞偲寢榑偝傟偨19)丅丂偡側傢偪丄乽俀庬椶埲忋偺僋儘乕僨傿儞偑嫟廳崌偟偰僿僥儘億儕儅乕偱偁傞俿俰僗僩儔儞僪傪宍惉偟丄岦偐偄崌偆嵶朎枌拞偺僗僩儔儞僪偳偆偟偑僋儘乕僨傿儞娫偺僿僥儘僼傿儕僢僋側愙拝偵傛傝懳崌偟偰丄嵶朎枌娫偺嫍棧傪僛儘偵偟偰偄傞偺偑丄俿俰偱偁傞乿偲寢榑偱偒傞丅丂偨偩偟丄幚嵺偵偼僆僋儖僨傿儞傕偙偺俿俰僗僩儔儞僪偵慻傒崬傑傟偰偍傝丄偝傜偵暋嶨側暘巕峔抸偑梊憐偝傟傞丅丂偳偺傛偆偵偙傟傜偺抈敀幙偑攝抲偝傟偰俿俰僗僩儔儞僪偑宍惉偝傟偰偄傞偐偼崱屻偺揹巕尠旝嬀側偳傪梡偄偨峔憿夝愅傪懸偨側偗傟偽側傜側偄丅
俈丏僶儕傾乕婡擻偲僋儘乕僨傿儞
丂俿俰僗僩儔儞僪偺婎杮峔憿偑僋儘乕僨傿儞偵傛偭偰峔惉偝傟偰偄傞偙偲偼柧傜偐偵側偭偨偑丄偦傟偱偼丄偙偺僋儘乕僨傿儞傛傝側傞俿俰僗僩儔儞僪偑忋旂嵶朎傗撪旂嵶朎僔乕僩偺僶儕傾乕婡擻傪幚嵺偵扴偭偰偄傞偺偱偁傠偆偐丠丂偁傞僋儘乕僨傿儞傪忋旂嵶朎偵夁忚敪尰偝偣偰傕丄俿俰僗僩儔儞僪偺悢乮俿俰偺敪払嬶崌乯偑丄偦傟偧傟偺忋旂嵶朎偱壗傜偐偺婡峔偱婯掕偝傟偰偄傞傜偟偔丄俿俰僗僩儔儞僪偺悢偑挊偟偔憹偊偰僶儕傾乕婡擻偑嬌抂偵忋徃偡傞偙偲偼側偄傛偆偱偁傞丅丂堦曽丄僋儘乕僨傿儞偺敪尰傪梷偊傞曽岦偺幚尡傕丄偦傟偧傟偺忋旂嵶朎偵婔庬椶傕偺僋儘乕僨傿儞偑敪尰偟偰偄傞偨傔偵堈偟偔側偄偑丄嵟嬤丄挵撪嵶嬠偺撆慺傪棙梡偡傞偲偄偆儐僯乕僋側曽朄偱丄僶儕傾乕婡擻偲僋儘乕僨傿儞偺娭學偑柧椖偵帵偝傟偨20)丅
丂忋弎偟偨傛偆偵丄僋儘乕僨傿儞亅侾偲亅俀偵帡偨暘巕傪僨乕僞儀乕僗偐傜扵偟偨帪偵丄挵撪嵶嬠乮Clostridium
perforingens乯偺儁僾僠僪惈撆慺偺儗僙僾僞乕乮俠俹俤亅俼乯21)傕丄僋儘乕僨傿儞僼傽儈儕乕偺堦堳偱偁傞偲偄偆偙偲偵婥晅偄偨乮尰嵼偱偼僋儘乕僨傿儞亅係偲屇偽傟偰偄傞乯丅丂偙偺撆慺偼丄俠枛抂敿暘偱儗僙僾僞乕偵寢崌偟丄俶枛抂敿暘偱嵶朎枌偵寠傪偁偗傞偲峫偊傜傟偰偄偨偺偱丄傑偢丄俠枛抂懁敿暘偵憡摉偡傞儁僾僠僪乮俠亅俠俹俤乯傪戝挵嬠偱嶌惢偟丄偙偺儁僾僠僪偑攟梴忋旂嵶朎偱偁傞俵俢俠俲嵶朎偺僶儕傾乕婡擻偵偳偺傛偆側塭嬁傪梌偊傞偐傪挷傋偨丅丂
 俵俢俠俲嵶朎偵偼丄偍傕偵丄僋儘乕僨傿儞亅侾偲亅係偑敪尰偟偰偄偨偑丄 俠亅俠俹俤偼僋儘乕僨傿儞亅侾偵偼寢崌偣偢丄僋儘乕僨傿儞亅係偵偩偗偵寢崌偡傞偙偲偑妋擣偝傟偨丅
嫽枴怺偄偙偲偵丄俵俢俠俲嵶朎傪俠亅俠俹俤懚嵼壓偱攟梴偡傞偲丄 侾帪娫埲撪偵偦偺俿俰偐傜僋儘乕僨傿儞亅係偺傒偑慖戰揑偵徚偊巒傔丄係帪娫埵偱僋儘乕僨傿儞亅侾偺傒偐傜側傞俿俰偵曄壔偟偨丅丂偙傟偵暯峴偟偰丄揹婥掞峈偲偟偰應掕偝傟傞僶儕傾乕婡擻偼嬌抂偵掅壓偟偨丅丂偙偺帪丄俿俰僗僩儔儞僪偼丄
俠亅俠俹俤偺懚嵼壓偱丄媫懍偵梟偗偨傛偆偵夡傟巒傔丄嵟廔揑偵僗僩儔儞僪偺悢偑敿暘埲壓偵尭傞偙偲偑暘偐偭偨乮恾俉乯丅丂偙偺儁僾僠僪傪愻偄棳偡偲丄嵞傃僋儘乕僨傿儞亅係偑俿俰偵擹弅偟巒傔丄僶儕傾乕婡擻傕夞暅偟偨丅丂埲忋偺寢壥偼丄僋儘乕僨傿儞偑俿俰僗僩儔儞僪傪宍惉偡傞偙偲偵傛傝丄忋旂嵶朎傗撪旂嵶朎僔乕僩偺僶儕傾乕婡擻傪捈愙扴偭偰偄傞偙偲傪弶傔偰帵偟偨傕偺偱偁傞丅
俵俢俠俲嵶朎偵偼丄偍傕偵丄僋儘乕僨傿儞亅侾偲亅係偑敪尰偟偰偄偨偑丄 俠亅俠俹俤偼僋儘乕僨傿儞亅侾偵偼寢崌偣偢丄僋儘乕僨傿儞亅係偵偩偗偵寢崌偡傞偙偲偑妋擣偝傟偨丅
嫽枴怺偄偙偲偵丄俵俢俠俲嵶朎傪俠亅俠俹俤懚嵼壓偱攟梴偡傞偲丄 侾帪娫埲撪偵偦偺俿俰偐傜僋儘乕僨傿儞亅係偺傒偑慖戰揑偵徚偊巒傔丄係帪娫埵偱僋儘乕僨傿儞亅侾偺傒偐傜側傞俿俰偵曄壔偟偨丅丂偙傟偵暯峴偟偰丄揹婥掞峈偲偟偰應掕偝傟傞僶儕傾乕婡擻偼嬌抂偵掅壓偟偨丅丂偙偺帪丄俿俰僗僩儔儞僪偼丄
俠亅俠俹俤偺懚嵼壓偱丄媫懍偵梟偗偨傛偆偵夡傟巒傔丄嵟廔揑偵僗僩儔儞僪偺悢偑敿暘埲壓偵尭傞偙偲偑暘偐偭偨乮恾俉乯丅丂偙偺儁僾僠僪傪愻偄棳偡偲丄嵞傃僋儘乕僨傿儞亅係偑俿俰偵擹弅偟巒傔丄僶儕傾乕婡擻傕夞暅偟偨丅丂埲忋偺寢壥偼丄僋儘乕僨傿儞偑俿俰僗僩儔儞僪傪宍惉偡傞偙偲偵傛傝丄忋旂嵶朎傗撪旂嵶朎僔乕僩偺僶儕傾乕婡擻傪捈愙扴偭偰偄傞偙偲傪弶傔偰帵偟偨傕偺偱偁傞丅
俉丏paracellular 僠儍僱儖偲僋儘乕僨傿儞
丂堎側傞僋儘乕僨傿儞娫偱僿僥儘僼傿儕僢僋側愙拝偑偱偒傞偲弎傋偨偑丄偙傟偼扨弮壔偟偡偓偰偄傞偙偲傕暘偐偭偰偒偨丅丂忋弎偟偨傛偆偵俠侾俴嵶朎偲俠俁俴嵶朎偺娫丄傑偨偼俠俀俴嵶朎偲俠俁俴嵶朎偺娫偵偼僿僥儘僼傿儕僢僋側愙拝偵傛傝棫攈側俿俰偑宍惉偝傟傞偑丄俠侾俴嵶朎偲俠俀俴嵶朎傪崿崌攟梴偟偰傕丄偦偺娫偵偼俿俰僗僩儔儞僪偑宍惉偝傟側偄偺偱偁傞19)丅丂偡側傢偪丄僋儘乕僨傿儞偺慻傒崌傢偣偵傛偭偰丄偦偺愙拝嫮搙偵柧傜偐側嵎偑偁傞傛偆偵傒偊傞丅丂偦偺傛偆側偙偲傪慜採偵偟偰丄儌僨儖俢傪挱傔偰傒傞偲丄偙偺俿俰偲偄偆峔憿偼側偐側偐暋嶨側僔僗僥儉偱偁傞偲偄偆偙偲偵婥晅偔丅丂椺偊偽丄僋儘乕僨傿儞乕侾偲僋儘乕僨傿儞乕俁偱儌僨儖俢偺傛偆側懳崌偡傞俀杮偺僗僩儔儞僪傪宍惉偟偰偄傞応崌偲丄僋儘乕僨傿儞乕侾偲僋儘乕僨傿儞乕俀偱宍惉偟偰偄傞応崌傪峫偊偰傒傛偆乮恾俋乯丅丂
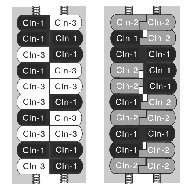 慜幰偱偼僋儘乕僨傿儞乕侾偲乕俁偺僿僥儘億儕儅乕偑懳崌偡傞偺偱丄僋儘乕僨傿儞乕侾偳偆偟偍傛傃僋儘乕僨傿儞乕俁偳偆偟偺儂儌僼傿儕僢僋側愙拝偲丄僋儘乕僨傿儞乕侾偲僋儘乕僨傿儞乕俁偺僿僥儘僼傿儕僢僋側愙拝偑崿嵼偡傞偑丄偄偢傟傕斾妑揑嫮偔愙拝偡傞丅丂偟偐偟丄屻幰偱偼丄僋儘乕僨傿儞乕侾偳偆偟偍傛傃僋儘乕僨傿儞乕俀偳偆偟偺儂儌僼傿儕僢僋側愙拝偼嫮偄偑丄嫮惂揑偵岦偐偄崌傢偝傟偨僋儘乕僨傿儞乕侾偲僋儘乕僨傿儞乕俀偼嫮偔偼愙拝偱偒側偄丅丂偟偨偑偭偰丄慜幰偺俿俰偺曽偑屻幰傛傝嫮偄僶儕傾乕婡擻傪帵偡偙偲偑婜懸偱偒傞丅丂偧傟偧傟扨撈偱傕俿俰僗僩儔儞僪傪宍惉偱偒傞侾俆庬椶傕偺僋儘乕僨傿儞偑懚嵼偡傞堄枴偼丄偙偺傛偆側偲偙傠偵偁傞偺偱偼側偄偩傠偆偐丠丂偦偺俿俰偵娷傑傟傞僋儘乕僨傿儞偺庬椶偲崿崌偺妱崌偵傛偭偰丄懳崌偡傞俿俰僗僩儔儞僪娫偵嶌傞乬寠乭偺悢偲惈幙傪曄偊偰丄俿俰偺僶儕傾乕婡擻傪岻傒偵挷愡偟偰偄傞偺偱偼側偄偩傠偆偐丠
慜幰偱偼僋儘乕僨傿儞乕侾偲乕俁偺僿僥儘億儕儅乕偑懳崌偡傞偺偱丄僋儘乕僨傿儞乕侾偳偆偟偍傛傃僋儘乕僨傿儞乕俁偳偆偟偺儂儌僼傿儕僢僋側愙拝偲丄僋儘乕僨傿儞乕侾偲僋儘乕僨傿儞乕俁偺僿僥儘僼傿儕僢僋側愙拝偑崿嵼偡傞偑丄偄偢傟傕斾妑揑嫮偔愙拝偡傞丅丂偟偐偟丄屻幰偱偼丄僋儘乕僨傿儞乕侾偳偆偟偍傛傃僋儘乕僨傿儞乕俀偳偆偟偺儂儌僼傿儕僢僋側愙拝偼嫮偄偑丄嫮惂揑偵岦偐偄崌傢偝傟偨僋儘乕僨傿儞乕侾偲僋儘乕僨傿儞乕俀偼嫮偔偼愙拝偱偒側偄丅丂偟偨偑偭偰丄慜幰偺俿俰偺曽偑屻幰傛傝嫮偄僶儕傾乕婡擻傪帵偡偙偲偑婜懸偱偒傞丅丂偧傟偧傟扨撈偱傕俿俰僗僩儔儞僪傪宍惉偱偒傞侾俆庬椶傕偺僋儘乕僨傿儞偑懚嵼偡傞堄枴偼丄偙偺傛偆側偲偙傠偵偁傞偺偱偼側偄偩傠偆偐丠丂偦偺俿俰偵娷傑傟傞僋儘乕僨傿儞偺庬椶偲崿崌偺妱崌偵傛偭偰丄懳崌偡傞俿俰僗僩儔儞僪娫偵嶌傞乬寠乭偺悢偲惈幙傪曄偊偰丄俿俰偺僶儕傾乕婡擻傪岻傒偵挷愡偟偰偄傞偺偱偼側偄偩傠偆偐丠
丂偙偺傛偆側壖愢偵傛偔崌抳偡傞帠幚偑丄偛偔嵟嬤丄忢愼怓懱楎惈堚揱宍幃傪偲傞掅儅僌僱僔僂儉寣徢偺尨場堚揱巕偺夝愅偐傜丄柧傜偐偵偝傟偨22)丅丂儅僌僱僔僂儉僀僆儞偼丄恡憻偺巺媴懱偱堦扷鄅夁偝傟丄擜拞偵弌偨屻偱丄偦偺戝晹暘偑僿儞儗偺忋峴媟偱嵞媧廂偝傟傞偑丄偙傟傜偺壠宯偺姵幰偱偼丄偙偺嵞媧廂偑慡偔峴傢傟側偄丅丂偦偺偨傔丄寣拞偺儅僌僱僔僂儉僀僆儞擹搙偑掅壓偡傞偲偄偆堚揱昦偱偁傞丅丂儅僌僱僔僂儉僀僆儞偺嵞媧廂偼丄忋峴媟偺忋旂偵偍偄偰丄transcellular
pathway偱偼側偔丄戝晹暘偑pracellular pathway傪夘偡傞庴摦揑側桝憲偵傛偭偰扴傢傟偰偄傞偙偲偑抦傜傟偰偄偨丅丂偦偺尨場堚揱巕偑億僕僔儑僫儖僋儘乕僯儞僌朄偵傛傝摨掕偝傟丄偦偺堚揱巕嶻暔偺傾儈僲巁攝楍偑悇掕偝傟偰傒傞偲丄嬃偄偨偙偲偵僋儘乕僨傿儞僼傽儈儕乕偵懏偡傞偙偲偑柧傜偐偵側偭偨偺偱偁傞丅丂偙偺僋儘乕僨傿儞偼丄偦傟傑偱摨掕偝傟偰偄偨侾俆庬椶偺僋儘乕僨傿儞偺偄偢傟偲傕堦抳偣偢丄僋儘乕僨傿儞乕侾俇乮堚揱巕傪摨掕偟偨僌儖乕僾偼paracellin-1
偲屇傫偱偄傞偑乯偲偄偆傋偒傕偺偱偁偭偨丅丂偙偺僋儘乕僨傿儞偼丄僿儞儗偺忋峴媟偩偗偵敪尰偟偰偄傞摿庩側僋儘乕僨傿儞偱丄偦偺敪尰偑側偔側傞偲乮傑偨偼婡擻偑梷惂偝傟傞偲乯paracellular
pathway傪夘偟偨儅僌僱僔僂儉僀僆儞偺桝憲偑慾奞偝傟傞偙偲偐傜丄偙偺僋儘乕僨傿儞偑俿俰僗僩儔儞僪偺拞偱丄儅僌僱僔僂儉僀僆儞傪斾妑揑慖戰揑偵捠偡僠儍僱儖乮寠乯傪宍惉偟偰偄傞偲峫偊傜傟偨丅丂偳偺傛偆偵偟偰偙偺僋儘乕僨傿儞偑僠儍僱儖傪宍惉偡傞偐偼崱屻偺栤戣偱偁傞偑丄忋弎偟偨傛偆偵丄傕偟丄偙偺僋儘乕僨傿儞偑僿儞儗偺忋峴媟偺忋旂嵶朎偵敪尰偟偰偄傞懠偺僋儘乕僨傿儞偲僿僥儘僼傿儕僢僋側愙拝傪偡傞忋偱憡惈偑埆偄偲壖掕偡傞偲丄懳崌偡傞俀杮偺俿俰僗僩儔儞僪娫偵僋儘乕僨傿儞乕侾俇乮paracellin-1)偲偦偺憡惈偺埆偄僋儘乕僨傿儞偲偵傛偭偰乬寠乭偑嶌傜傟丄偦偙傪儅僌僱僔僂儉僀僆儞偑捠傞偙偲偼梕堈偵憐憸偱偒傞丅丂偙偺傛偆側敪尒偵傛傝丄屆偔偐傜偦偺懚嵼偑梊憐偝傟偰偄偨paracellular
僠儍僱儖偺幚懱偑丄僋儘乕僨傿儞偦偺傕偺偱偁傞偙偲偑帵嵈偝傟偨丅
俋丏僞僀僩僕儍儞僋僔儑儞尋媶偺崱屻丗傑偲傔偵偐偊偰
丂偙偺憤愢偱偼丄変乆偺尋媶偺棳傟傪捛偭偰丄俿俰偺愙拝暘巕丄偡側傢偪俿俰僗僩儔儞僪傪宍惉偡傞撪嵼惈枌抈敀幙偵偮偄偰丄尰嵼傑偱偵柧傜偐偵側偭偨偙偲傪傑偲傔偰傒偨丅丂嵶朎娫愙拝偲偄偆尰徾偑暘巕偺尵梩偱岅傜傟傞傛偆偵側偭偰偐傜媣偟偄偑丄俿俰偲偄偆摿庩側嵶朎娫愙拝傪巌傞暘巕偺摨掕偼抶傟偰偄偨丅丂僋儘乕僨傿儞暘巕偺偐側傝偺晹暘偑嵶朎枌撪偵杽傑偭偰偄傞偨傔偵丄峈懱偵傛傞暘巕偺摨掕傗婡擻慾奞偑擄偟偐偭偨偙偲傕堦場偱偁偭偨傛偆偵巚傢傟傞丅丂僋儘乕僨傿儞暘巕孮偼丄敪尒偝傟偰偐傜擔偑愺偄偨傔偵丄傑偩晄柧側揰偑懡偄丅丂偟偐偟丄俿俰僗僩儔儞僪偺婎杮揑側暘巕峔抸偑柧傜偐偵偝傟偨崱丄俿俰偺尋媶偼偄偔偮偐偺曽岦偵戝偒偔敪揥偡傞傕偺偲婜懸偝傟傞丅丂崱屻偺敪揥偺曽岦傪媍榑偡傞偙偲偱丄偙偺憤愢偺傑偲傔偲偟偨偄丅
丂傑偢丄戞堦偵丄杮憤愢偱徻偟偔弎傋偰偒偨傛偆偵丄俿俰偺僶儕傾婡擻偺暘巕儗儀儖偱偺棟夝偑怺傑傞偱偁傠偆丅丂俿俰偺僶儕傾婡擻丄偍傛傃偦偺挷愡婡擻偑丄懡嵶朎惗暔偺屄懱宍惉偺忋偱偳偺傛偆偵廳梫側栶妱傪壥偨偟偰偄傞偐偑丄偦傟偧傟偺僋儘乕僨傿儞堚揱巕偺僲僢僋傾僂僩儅僂僗傪嶌惢偡傞偙偲偵傛傝丄柧傜偐偵偝傟傞偱偁傠偆丅
偦偟偰丄僶儕傾乕婡擻偺堎忢偲偝傑偞傑側幘姵丒昦懺偲偺娭學傕柧傜偐偵側傞偱偁傠偆丅丂偝傜偵丄Clostridium
perforingens偺撆慺偺幚尡偱傕柧傜偐側傛偆偵丄俿俰偺僶儕傾乕婡擻傪恖堊揑偵憖嶌偡傞曽朄偑妋棫偝傟丄擼寣娗娭栧側偳傪堦帪揑偵攋夡偡傞偙偲偑壜擻偲側傝丄僪儔僢僌僨儕僶儕乕朄偺奐敪偺忋偱傕戝偒側恑曕傪傕偨傜偡偱偁傠偆丅丂偡側傢偪丄偙傟傑偱偺transcellular
pathway偺惗棟妛偵懳偟偰丄慡偔怴偟偄paracellular pathway偺惗棟妛偺敪揥偲偦偺墳梡偑婜懸偱偒傞丅
丂戞擇偵丄偙偙偱偼慡偔怗傟側偐偭偨偙偲偱偁傞偑丄忋旂嵶朎傗撪旂嵶朎偺嬌惈宍惉偵偍偗傞俿俰偺栶妱偺棟夝傕暘巕儗儀儖偱怺傑傞偱偁傠偆丅丂俿俰僗僩儔儞僪偼丄傾僺僇儖枌僪儊僀儞偲僶僜儔僥儔儖枌僪儊僀儞偺娫偵丄僆僀儖僼僃儞僗偺傛偆偵挘傝弰傜偝傟偰偄傞偨傔丄擇偮偺僪儊僀儞傪暘偗傞栶妱乮僼僃儞僗婡擻乯傪壥偨偟偰偄傞偲憐掕偝傟偰偒偨2)丅丂偙偺憐掕偦偺傕偺偑惓偟偄偐偳偆偐丄僋儘乕僨傿儞偑摨掕偝傟偨崱丄幚尡揑偵専徹偱偒傞偱偁傠偆丅丂傑偨丄俿俰僗僩儔儞僪偺嵶朎幙懁偵偼丄僆僋儖僨傿儞丒僋儘乕僨傿儞偲捈愙寢崌偡傞倅俷亅侾丄倅俷亅俀丄倅俷亅俁側偳偺棤懪偪抈敀幙偺懠偵23,24)丄庬乆偺忋旂嵶朎偺宍懺宍惉僔僌僫儖傪扴偆偲巚傢傟傞場巕偑廤愊偟偰偄傞偙偲偑嵟嬤柧傜偐偵側傝偮偮偁傞25)丅丂偙偺傛偆側僔僌僫儖揱払偺僙儞僞乕偺堦偮偲偟偰偺俿俰偺栶妱偺夝愅傕崱屻偺廳梫側曽岦偱偁傠偆丅
丂堦曽偱丄僆僋儖僨傿儞偺婡擻偼埶慠偲偟偰撲偵曪傑傟偰偄傞丅丂僆僋儖僨傿儞僲僢僋傾僂僩儅僂僗偼丄堦尒惓忢偵惗傑傟傞偑丄偦偺屻偺惉挿偑抶偔丄堓丄懥塼態丄擼丄崪側偳偵偦傟偧傟摿挜揑側昦曄傪惗偠丄巕懛傪巆偡偙偲偑偱偒側偄乮惸摗傜丄搳峞弨旛拞乯丅丂偙偺儅僂僗偼丄
傑偩変乆偑慡偔抦傜側偄廳梫側栶妱傪俿俰偑壥偨偟偰偄傞偙偲傪嫮偔帵嵈偟偰偄傞丅丂俿俰偺廃曈偵偼丄傑偩傑偩丄懡偔偺撲夝偒偺妝偟傒偑巆偭偰偄傞丅
丂偙偺尋媶偼丄搒椪彴尋丒崙棫惗棟尋埲棃偺拠娫偱偁傝丄尰嵼偺変乆偺尋媶幒偺僗僞僢僼偱偁傞寧揷憗抭巕丄暷懞廳怣丄塱熀徍椙丄埳摗夒旻偺奺攷巑偑恑傔偰偒偨嵶朎娫愙拝憰抲偺暘巕惗暔妛揑尋媶偺墑挿慄忋偵丄奺攷巑偺嫤椡偵傛傝惗傑傟偰偒偨傕偺偱偁傝丄偙偙偵怺偔姶幱偟傑偡丅丂傑偨丄偙偙偵徯夘偟偨尋媶偺懡偔偼丄戝妛堾惗偵傛傝恑傔傜傟偨傕偺偱偁傞偙偲傪婰偟丄姶幱偺堄傪昞偟傑偡丅
乮曗乯杮暥傪彂偒忋偘偰偐傜報嶞傑偱偺娫偵丄僋儘乕僨傿儞亅侾侾偺僲僢僋傾僂僩儅僂僗偑曬崘偝傟偨丂[Grow,A., Southwood,C.M., Li,J.S., Pariali,M., Riordan,G.P., Brodie,S.E., Danias,J., Bronstein,J.M., Kachar,B. & Lazzarinin,R.A. (1999) Cell 99,649-659]丅丂僋儘乕僨傿儞亅侾侾偼俷俽俹乮oligodendrocyte specific protein乯偲偟偰侾俋俋6擭偵堚揱巕偲偟偰曬崘偝傟偰偄偨傕偺偱偁傞偑丄偙偺堚揱巕嶻暔偑俿俰偺峔惉場巕偱偁傞偙偲偑暘偐傞埲慜偐傜丄偙偺堚揱巕偺婡擻夝愅偺堦娐偲偟偰傾儊儕僇偺僌儖乕僾偵傛傝僲僢僋傾僂僩儅僂僗偺嶌惢偑恑傔傜傟偰偄偨傜偟偄丅丂杮暥拞偱弎傋偨傛偆偵丄偙偺僋儘乕僨傿儞偼拞悤恄宱偺儈僄儕儞偲惛憙偺僙儖僩儕嵶朎偺俿俰偵摿堎揑偵懚嵼偟偰偄傞摿庩側傕偺偱偁傞偙偲傪変乆偼曬崘偟偰偄偨偑18乯丄梊憐捠傝偙偺儅僂僗偱偼偙傟傜偺嵶朎偐傜俿俰偑徚偊偰偍傝丄桳悜恄宱偺揱摫懍搙偺掅壓偲惛巕宍惉晄慡偑娤嶡偝傟偨丅丂偙傟偑僋儘乕僨傿儞僼傽儈儕乕偺儊儞僶乕偺僲僢僋傾僂僩儅僂僗偲偟偰偼嵟弶偺曬崘偱偁傝丄僋儘乕僨傿儞偺廳梫惈偑屄懱儗儀儖偱徹柧偝傟偨偙偲偵側傞丅丂傑偨丄俀侽侽侽擭侾寧偺抜奒偱丄僋儘乕僨傿儞偼侾俉庬椶傑偱摨掕偝傟偰偄傞丅
暥丂專
1) Claude, P. (1978) J. Membr. Biol. 10, 219-232
2) Gumbiner, B. (1993) J. Cell Biol. 123, 1631-1633
3) Farquhar, M.G., & Palade, G.E. (1963) J. Cell Biol.
17, 375-412
4) Staehelin, L.A. (1974) Int. Rev. Cytol. 39, 191-283
5) Pinto da Silva, P., & Kachar, B. (1982) Cell 28,
441-450
6) Tsukita, Sh., & Tsukita, Sa. (1989) J. Cell Biol. 108,
31-41
7) Itoh, M., Nagafuchi, A., Yonemura, S., Kitani-Yasuda, T., Tsukita,
Sa., & Tsukita, Sh. (1993) J. Cell Biol. 121,
491-502
8) Furuse, M., Hirase, T., Itoh, M., Nagafuchi, A., Yonemura,
S., Tsukita, Sa., & Tsukita, Sh. (1993) J.Cell Biol.
123, 1777-1788
9) Ando-Akatsuka, Y., Saitou, M., Hirase, T., Kishi, M., Sakakibara,
A., Itoh, M., Yonemura, S., Furuse, M., & Tsukita, Sh. (1996)
J. Cell Biol. 133, 43-47
10) Matter, K., & Balda, M. (1999) Int. Rev. Cytol. 186,
117-146
11) Saitou, M., Fujimoto, K., Doi, Y., Itoh, M., Fujimoto, T.,
Furuse, M., Takano, H., Noda, T., & Tsukita. Sh. (1998) J.Cell
Biol. 141, 397-408
12) Furuse, M., Fujita, K., Hiiragi, T., Fujimoto, K., & Tsukita,
Sh. (1998) J. Cell Biol. 141, 1539-1550
13) Kubota, K., Furuse, M., Sasaki, H., Sonoda, N., Fujita, K.,
Nagafuchi, A., & Tsukita, Sh. (1999) Curr Biol. 9,
1035-1038
14) Furuse, M., Sasaki, H., Fujimoto, K., & Tsukita, Sh. (1998)
J. Cell Biol. 143, 391-401
15) Morita, K., Furuse, M., Fujimoto, K., & Tsukita, Sh. (1999)
Proc. Natl. Acad. Sci. USA 96, 511-516
16) Tsukita, Sh., & Furuse, M. (1999) Trend. Cell Biol.
9, 268-273
17) Morita, K., Sasaki, H., Furuse, M., & Tsukita, Sh. (1999)
J.Cell Biol. 147, 185-194
18) Morita, K., Sasaki, H., Fujimoto, K., Furuse, M., & Tsukita.
Sh. (1999) J. Cell Biol. 145, 579-788
19) Furuse, M., Sasaki, H., & Tsukita, Sh. (1999) J. Cell
Biol. in press.
20) Sonoda, N., Furuse, M., Sasaki, H., Yonemura, S., Katahira,
J., Horiguchi, Y., & Tsukita, Sh. (1999) J. Cell Biol.
147, 195-204
21) Katahira, J., Inoue, N., Horiguchi, Y., Matsuda, M., &
Sugimoto, N. (1997) J. Cell Biol. 136, 1239-1247
22) Simon, D.B., Lu, Y., Choate, K.A., Velazquez, H., Al-Sabban,
E., Praga, M., Casari, G., Bettinelli, A., Colussi, G., Rodriguez-Soriano,
J., McCredie, D., Milford, D., Sanjad, & S., Lifton, R.P.
(1999) Science 285, 103-106
23) Furuse, M., Itoh, M., Hirase, T., Nagafuchi, A., Yonemura,
S., Tsukita, Sa., and Tsukita, Sh. (1994) J. Cell Biol. 127,
1617-1626
24) Itoh, M., Furuse, M., Saitou, M., Morita, K., Kubota, K.,
and Tsukita, Sh. (1999) J. Cell Biol. in press
25) Tsukita, Sh., Furuse, M., & Itoh, M. (1999) Curr. Opin.
Cell Biol. 11, 628-633
![]()