僋儘乕僨傿儞偑慄堐夎嵶朎偵僞僀僩僕儍儞僋僔儑儞傪嶌偭偨両乮侾俋俋俋擭侾寧侾擔偺婰帠乯
乣僼僃僀儞僩傪偐偗偪傖偭偰偛柶側偝偄乣
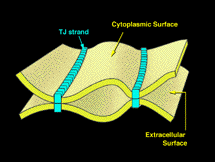 嵶朎娫偺暔幙偺楻傟傪杊偖暘巕婡峔傪巌傞偺偼丄屆偔偐傜宍懺妛揑偵抦傜傟偰偄傞僞僀僩僕儍儞僋僔儑儞偱偁傞丅丂僞僀僩僕儍儞僋僔儑儞偼嵍恾偺傛偆偵丄撪嵼惈枌抈敀幙偑枌撪偱慄忬偵暲傫偩億儕儅乕乮僗僩儔儞僪乯傪宍惉偡傞偙偲偵傛傝丄偙偺僶儕傾乕婡擻傪壥偨偟偰偄傞偲峫偊傜傟偰偒偨丅丂偙偺僞僀僩僕儍儞僋僔儑儞偺僗僩儔儞僪偱摥偔愙拝暘巕傪尒偮偗傟偽丄懡嵶朎惗暔偺宍懺宍惉偺暘巕婡峔傪棟夝偡傞忋偱傕丄傑偨丄庬乆偺昦懺傪棟夝偡傞忋偱傕丄戝偒側恑曕偱偁傞偙偲偼丄屆偔偐傜擣幆偝傟偰偒偨丅丂偙偺傛偆側忬嫷壓偱丄変乆偼僆僋儖僨傿儞傪侾俋俋俁擭偺屻敿偵弶傔偰摨掕偡傞偙偲偵惉岟偟偨丅丂偙偺帪偺斀嬁偼乮巆擮側偑傜擔杮崙撪偐傜偺斀嬁偼彮側偐偭偨偑丄悽奅偐傜偺斀嬁偼乯偡偽傜偟偔戝偒偐偭偨丅丂偦偟偰丄懡偔偺尋媶幒偑僆僋儖僨傿儞偺夝愅傪巒傔丄僆僋儖僨傿儞偑僞僀僩僕儍儞僋僔儑儞偺婡擻慺巕偱偁傞偲偡傞榑暥偑師乆偲弌偝傟偨丅
嵶朎娫偺暔幙偺楻傟傪杊偖暘巕婡峔傪巌傞偺偼丄屆偔偐傜宍懺妛揑偵抦傜傟偰偄傞僞僀僩僕儍儞僋僔儑儞偱偁傞丅丂僞僀僩僕儍儞僋僔儑儞偼嵍恾偺傛偆偵丄撪嵼惈枌抈敀幙偑枌撪偱慄忬偵暲傫偩億儕儅乕乮僗僩儔儞僪乯傪宍惉偡傞偙偲偵傛傝丄偙偺僶儕傾乕婡擻傪壥偨偟偰偄傞偲峫偊傜傟偰偒偨丅丂偙偺僞僀僩僕儍儞僋僔儑儞偺僗僩儔儞僪偱摥偔愙拝暘巕傪尒偮偗傟偽丄懡嵶朎惗暔偺宍懺宍惉偺暘巕婡峔傪棟夝偡傞忋偱傕丄傑偨丄庬乆偺昦懺傪棟夝偡傞忋偱傕丄戝偒側恑曕偱偁傞偙偲偼丄屆偔偐傜擣幆偝傟偰偒偨丅丂偙偺傛偆側忬嫷壓偱丄変乆偼僆僋儖僨傿儞傪侾俋俋俁擭偺屻敿偵弶傔偰摨掕偡傞偙偲偵惉岟偟偨丅丂偙偺帪偺斀嬁偼乮巆擮側偑傜擔杮崙撪偐傜偺斀嬁偼彮側偐偭偨偑丄悽奅偐傜偺斀嬁偼乯偡偽傜偟偔戝偒偐偭偨丅丂偦偟偰丄懡偔偺尋媶幒偑僆僋儖僨傿儞偺夝愅傪巒傔丄僆僋儖僨傿儞偑僞僀僩僕儍儞僋僔儑儞偺婡擻慺巕偱偁傞偲偡傞榑暥偑師乆偲弌偝傟偨丅
偟偐偟丄変乆偼庍慠偲偟側偄姶怗傪帩偭偰偄偨丅丂変乆偼僆僋儖僨傿儞傪敪尒偟偰埲棃丄偦偺婡擻夝愅傪偟傛偆偲丄偄傠偄傠側偙偲傪帋傒偨偑丄懠偺尋媶幒偐傜弌偰偔傞傛偆側僋儕傾乕側僨乕僞傪摼傞偙偲偼偱偒側偐偭偨丅丂壗偐曄偩偧偲忢偵巚偭偰偄偨丅丂変乆偼丄僆僋儖僨傿儞偺儅僂僗儂儌儘僌傪弶傔偰僋儘乕僯儞僌偵惉岟偟偨帪丄偡偖偵丄娻尋偺栰揷揘惗攷巑偲嫟摨偱丄僆僋儖僨傿儞偺僲僢僋傾僂僩傪帋傒偨丅丂偦偺寢壥丄嵟怴榑暥偺崁偵偁傞傛偆偵丄僆僋儖僨傿儞偺側偄忋旂嵶朎偱傕丄棫攈側僞僀僩僕儍儞僋僔儑儞丒僗僩儔儞僪偑偁傞偙偲偑尒弌偝傟偨丅丂変乆偲偟偰偼丄杮摉偵徟偭偨丅丂偙偺僲僢僋傾僂僩偺寢壥偼丄僆僋儖僨傿儞埲奜偵僞僀僩僕儍儞僋僔儑儞丒僗僩儔儞僪傪偮偔傞枌撪嵼惈抈敀幙偑懚嵼偡傞偙偲傪柧椖偵帵偟偰偄傞丅丂偱傕丄僨乕僞儀乕僗偱専嶕偡傞尷傝丄僆僋儖僨傿儞偵偄傢備傞傾僀僜僞僀僾偑偁傞偲偼巚偊側偄丅丂偱偼丄壗偑僞僀僩僕儍儞僋僔儑儞傪嶌偭偰偄傞偺偐丠
僞僀僩僕儍儞僋僔儑儞傪宍惉偡傞撪嵼惈抈敀幙偺摨掕偼丄偙傟傑偱嵟傕擄偟偄偲偝傟偰偄偨暘栰偱偁傞丅丂偟偐偟丄崱偼丄僆僋儖僨傿儞偑偁傞丅丂僆僋儖僨傿儞偵寢崌偡傞抈敀幙傪扵偣偽傛偄偺偱偼側偄偐丅丂傕偪傠傫丄傑偢丄yeast
two-hybrid傪帋傒偨丅丂偙偙偱庢傟傞傕偺側傜丄変乆偺尋媶幒埲奜偐傜婛偵曬崘偑偁偭偨偱偁傠偆丅丂妋偐偵丄僱僈僥傿僽側寢壥偺傒摼傜傟偨丅丂俼俿亅俹俠俼傪巒傔丄柶塽捑崀丄偄傠偄傠傗偭偨丅丂偡傋偰僱僈僥傿僽僨乕僞偱偁偭偨丅丂偦偙偱丄嵟弶偺僕儍儞僋僔儑儞暘夋偵栠偭偨丅丂敪憐偼娙扨偱偁傞丅丂栚巜偡撪嵼惈枌抈敀幙偼丄僆僋儖僨傿儞偲摨條偵僕儍儞僋僔儑儞暘夋偵擹弅偟偰偄傞敜偱偁傞丅丂偦偺検傕丄僆僋儖僨傿儞偵懟怓側偄敜偱偁傞丅丂変乆偼丄偙偺傛偆側敪憐偐傜丄僕儍儞僋僔儑儞暘夋傪傕偆堦搙丄揙掙揑偵夝愅偟丄暘巕検俀俀倠俢偺係夞枌娧捠抈敀幙偑僞僀僩僕儍儞僋僔儑儞偵擹弅偡傞偙偲傪尒弌偟偨丅
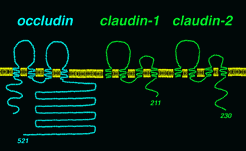 偙偺枌抈敀幙傪僋儘乕僨傿儞乮claudin)乕侾偲乕俀偲柤晅偗偨偑丄偦偺屻偺尋媶偱丄僋儘乕僨傿儞偼乕俉傑偱尒偮偐偭偰偄傞丅丂嵍恾偵偁傞傛偆偵丄彮側偔偲傕娞憻偺僞僀僩僕儍儞僋僔儑儞偺僗僩儔儞僪偼丄僆僋儖僨傿儞偲丄僋儘乕僨傿儞乕侾丄乕俀偺俁庬椶偺係夞枌娧捠抈敀幙偐傜弌棃偰偄傞偙偲偵側傞丅
偙偺枌抈敀幙傪僋儘乕僨傿儞乮claudin)乕侾偲乕俀偲柤晅偗偨偑丄偦偺屻偺尋媶偱丄僋儘乕僨傿儞偼乕俉傑偱尒偮偐偭偰偄傞丅丂嵍恾偵偁傞傛偆偵丄彮側偔偲傕娞憻偺僞僀僩僕儍儞僋僔儑儞偺僗僩儔儞僪偼丄僆僋儖僨傿儞偲丄僋儘乕僨傿儞乕侾丄乕俀偺俁庬椶偺係夞枌娧捠抈敀幙偐傜弌棃偰偄傞偙偲偵側傞丅
偦傟偱偼丄僋儘乕僨傿儞偲僆僋儖僨傿儞偼偳偺傛偆偵僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪偺宍惉傗丄僞僀僩僕儍儞僋僔儑儞偺僶儕傾乕婡擻丄僼僃儞僗婡擻偵峷專偟偰偄傞偺偱偁傠偆偐丠丂偙偺媈栤偵摎偊傞幚尡偼巒傑偭偨偽偐傝偱偁傞丅丂偟偐偟丄嵟嬤丄偙偺媈栤偺堦晹偵摎偊傞偒傢傔偰嫽枴怺偄寢壥傪摼偨丅丂徻嵶偼丄嵟怴偺榑暥偺崁偵徯夘偟偰偄傞偑丄僞僀僩僕儍儞僋僔儑儞傪帩偨側偄儅僂僗俴慄堐夎嵶朎偵僋儘乕僨傿儞乕侾傑偨偼僋儘乕僨傿儞乕俀偺們俢俶俙傪摫擖偟偨偲偙傠丄側傫偲丄摼傜傟偨僩儔儞僗僼僃僋僞儞僩娫偵棫攈側僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪偺僱僢僩儚乕僋偑偱偒偰偟傑偭偨偺偱偁傞丅丂偙偺峔憿傪丄捠忢偺搥寢妱抐儗僾儕僇朄偱娤嶡偟偨偺偑嵍壓恾偱丄杮島嵗偺嵟弶偺儂乕儉儁乕僕偵宖嵹偝傟偰偄偨傕偺偱偁傞丅丂偙偺僞僀僩僕儍儞僋僔儑儞僱僢僩儚乕僋偼嫄戝側傕偺偱丄岝妛尠旝嬀儗儀儖偱傕娤嶡偱偒傞丅丂偱偼丄僆僋儖僨傿儞偼偳偆偐丠丂偙傟傕徻嵶偼榑暥傪嶲徠偝傟偨偄偑丄寢榑偲偟偰丄僆僋儖僨傿儞偼僋儘乕僨傿儞偲嵶朎枌偺拞偱嫟廳崌偟偰僗僩儔儞僪傪嶌偭偰偄傞偙偲偑柧傜偐偵側偭偨丅
 懡嵶朎惗暔偺峔抸偺拞偱丄僞僀僩僕儍儞僋僔儑儞偺壥偨偡栶妱偼杮幙揑偵廳梫偱偁傞丅丂偦偺僞僀僩僕儍儞僋僔儑儞偺暘巕峔抸偑傎傏夝柧偝傟偨丅丂嵶朎惗暔妛傪昗炘偟偰偒偨尋媶幒偲偟偰偼丄偙傟偼偐側傝屩偭偰傕椙偄惉壥偱偼側偄偐偲棪捈偵巚偆丅丂偦傟傕丄崱偺帪戙偵偼捒偟偔丄傎傏偙偙傑偱撈憱偟偰偙傟偨偺偩偐傜丅丂側偤偙偺悽抦恏偄悽偺拞偱撈憱偱偒偨偺偐丠丂偙傟偼偐側傝塣偑椙偐偭偨偲巚偆丅丂偩偄偨偄丄僋儘乕僨傿儞偲偄偆杮柦偑戝晹暘嵶朎枌偺拞偵杽傑偭偰偄傞抈敀幙偱丄崱偺暘巕嵶朎惗暔妛偺媄弍偱曔傑偊擄偐偭偨偙偲偑丄偦傕偦傕偺嫮塣偺巒傑傝偱偁傞丅丂儌僲僋儘乕僫儖峈懱朄偱峈懱偑扤傕庢傟側偐偭偨偟丄嵶朎枌撪偺寢崌偼yeast
two-hybrid偑嬯庤偲偟偰偄偨丅丂偟偐偟丄崱偺忬嫷偱堦斣偍傕偟傠偄嫮塣偼丄僆僋儖僨傿儞偑杮柦偱側偐偭偨偙偲偱偁傠偆乮僆僋儖僨傿儞帺懱偼偍傕偟傠偔丄偦偺僲僢僋傾僂僩儅僂僗偺夝愅傕恑傒偮偮偁傞偑丄僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪宍惉偲尵偆揰偱偼杮柦偱偼側偄乯丅丂僆僋儖僨傿儞傪曬崘偟偰偐傜丄忋弎偟偨傛偆偵懡偔偺尋媶幒偑旘傃偮偄偨丅丂偦偺惉壥偑崱崰憡師偄偱曬崘偝傟偰偄傞乮侾俋俋俉擭搙偱崱傑偱偵俆侽埲忋偺榑暥偑宖嵹偝傟偰偄傞傛偆偱偁傞乯丅丂僆僋儖僨傿儞偵娭偡傞憤愢傑偱俀丆俁彂偐傟偰偄傞忬懺偵傑偱側偭偨丅丂偲偙傠偑丄宍偲偟偰偼丄崱擭偺慜敿偵丄変乆帺恎偑僆僋儖僨傿儞傪僲僢僋傾僂僩偟偰傕僞僀僩僕儍儞僋僔儑儞偼偱偒傞偲偄偆丄僽僠僐儚僔榑暥傪曬崘偟丄偝傜偵懅偮偖壣傕側偔俀偮乮幚嵺偵偼俁偮偵側傞偑乯偺僋儘乕僨傿儞榑暥傪敪昞偟偨乮乽懅偮偖壣偑側偄乿傛偆偵側偭偨偺偼丄変乆偲偟偰傕怴偟偄僞僀僩僕儍儞僋僔儑儞枌抈敀幙偺庤妡偐傝傪摼偢偟偰丄僽僠僐儚僔榑暥傪彂偔桬婥偑側偐偭偨偙偲偵傕傛偭偰偄傞偺偱丄埆婥偑偁偭偨栿偱偼側偄乯丅丂寢壥偲偟偰偼丄懡偔偺尋媶幒偵丄僒僢僇乕偱偄偆偲偙傠偺僼僃僀儞僩傪偐偗偨傛偆側忬嫷偑惗傑傟偰偟傑偭偨丅丂懡偔偺億僣僪僋傗妛惗偑僆僋儖僨傿儞偑僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪偺拞怱揑抈敀幙偩偲偄偆壖掕偺傕偲偵夝愅傪懕偗偰偄偨敜偱偁傞偑丄崱傗丄僋儘乕僨傿儞側偟偵偼僞僀僩僕儍儞僋僔儑儞偑岅傟側偄偱偁傠偆偲偄偆婏柇側忬嫷偑惗傑傟偨栿偱偁傞丅丂偙偺恖偨偪偵偼婥偺撆偩偑丄堦帪偺惷庘傪偱偒傞偩偗棙梡偟偰丄儕乕僪傪峀偘偨偄偺偑杮壒偱偁傞丅丂僞僀僩僕儍儞僋僔儑儞偼丄堦斒偵巚傢傟偰偄傞傛傝傕偐側傝杮幙揑側栤戣傪娷傫偱偄傞偺偱丄懡偔偺恖偑懺惃傪棫偰捈偟偰捛偭偐偗偰偔傞偙偲偼昁帄偱偁傞丅丂偙傟偼僆僋儖僨傿儞偺帪偵捝姶偟偨丅丂僂乕儉丄偙傟偐傜壗偵廤拞偟偰傗偭偰偄偔偐丠丂僲僢僋傾僂僩偐丠丂偦傟偲傕亅亅亅亅丅丂枅擔丄媍榑傪偟偰歑偭偰偄傞丅
懡嵶朎惗暔偺峔抸偺拞偱丄僞僀僩僕儍儞僋僔儑儞偺壥偨偡栶妱偼杮幙揑偵廳梫偱偁傞丅丂偦偺僞僀僩僕儍儞僋僔儑儞偺暘巕峔抸偑傎傏夝柧偝傟偨丅丂嵶朎惗暔妛傪昗炘偟偰偒偨尋媶幒偲偟偰偼丄偙傟偼偐側傝屩偭偰傕椙偄惉壥偱偼側偄偐偲棪捈偵巚偆丅丂偦傟傕丄崱偺帪戙偵偼捒偟偔丄傎傏偙偙傑偱撈憱偟偰偙傟偨偺偩偐傜丅丂側偤偙偺悽抦恏偄悽偺拞偱撈憱偱偒偨偺偐丠丂偙傟偼偐側傝塣偑椙偐偭偨偲巚偆丅丂偩偄偨偄丄僋儘乕僨傿儞偲偄偆杮柦偑戝晹暘嵶朎枌偺拞偵杽傑偭偰偄傞抈敀幙偱丄崱偺暘巕嵶朎惗暔妛偺媄弍偱曔傑偊擄偐偭偨偙偲偑丄偦傕偦傕偺嫮塣偺巒傑傝偱偁傞丅丂儌僲僋儘乕僫儖峈懱朄偱峈懱偑扤傕庢傟側偐偭偨偟丄嵶朎枌撪偺寢崌偼yeast
two-hybrid偑嬯庤偲偟偰偄偨丅丂偟偐偟丄崱偺忬嫷偱堦斣偍傕偟傠偄嫮塣偼丄僆僋儖僨傿儞偑杮柦偱側偐偭偨偙偲偱偁傠偆乮僆僋儖僨傿儞帺懱偼偍傕偟傠偔丄偦偺僲僢僋傾僂僩儅僂僗偺夝愅傕恑傒偮偮偁傞偑丄僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪宍惉偲尵偆揰偱偼杮柦偱偼側偄乯丅丂僆僋儖僨傿儞傪曬崘偟偰偐傜丄忋弎偟偨傛偆偵懡偔偺尋媶幒偑旘傃偮偄偨丅丂偦偺惉壥偑崱崰憡師偄偱曬崘偝傟偰偄傞乮侾俋俋俉擭搙偱崱傑偱偵俆侽埲忋偺榑暥偑宖嵹偝傟偰偄傞傛偆偱偁傞乯丅丂僆僋儖僨傿儞偵娭偡傞憤愢傑偱俀丆俁彂偐傟偰偄傞忬懺偵傑偱側偭偨丅丂偲偙傠偑丄宍偲偟偰偼丄崱擭偺慜敿偵丄変乆帺恎偑僆僋儖僨傿儞傪僲僢僋傾僂僩偟偰傕僞僀僩僕儍儞僋僔儑儞偼偱偒傞偲偄偆丄僽僠僐儚僔榑暥傪曬崘偟丄偝傜偵懅偮偖壣傕側偔俀偮乮幚嵺偵偼俁偮偵側傞偑乯偺僋儘乕僨傿儞榑暥傪敪昞偟偨乮乽懅偮偖壣偑側偄乿傛偆偵側偭偨偺偼丄変乆偲偟偰傕怴偟偄僞僀僩僕儍儞僋僔儑儞枌抈敀幙偺庤妡偐傝傪摼偢偟偰丄僽僠僐儚僔榑暥傪彂偔桬婥偑側偐偭偨偙偲偵傕傛偭偰偄傞偺偱丄埆婥偑偁偭偨栿偱偼側偄乯丅丂寢壥偲偟偰偼丄懡偔偺尋媶幒偵丄僒僢僇乕偱偄偆偲偙傠偺僼僃僀儞僩傪偐偗偨傛偆側忬嫷偑惗傑傟偰偟傑偭偨丅丂懡偔偺億僣僪僋傗妛惗偑僆僋儖僨傿儞偑僞僀僩僕儍儞僋僔儑儞僗僩儔儞僪偺拞怱揑抈敀幙偩偲偄偆壖掕偺傕偲偵夝愅傪懕偗偰偄偨敜偱偁傞偑丄崱傗丄僋儘乕僨傿儞側偟偵偼僞僀僩僕儍儞僋僔儑儞偑岅傟側偄偱偁傠偆偲偄偆婏柇側忬嫷偑惗傑傟偨栿偱偁傞丅丂偙偺恖偨偪偵偼婥偺撆偩偑丄堦帪偺惷庘傪偱偒傞偩偗棙梡偟偰丄儕乕僪傪峀偘偨偄偺偑杮壒偱偁傞丅丂僞僀僩僕儍儞僋僔儑儞偼丄堦斒偵巚傢傟偰偄傞傛傝傕偐側傝杮幙揑側栤戣傪娷傫偱偄傞偺偱丄懡偔偺恖偑懺惃傪棫偰捈偟偰捛偭偐偗偰偔傞偙偲偼昁帄偱偁傞丅丂偙傟偼僆僋儖僨傿儞偺帪偵捝姶偟偨丅丂僂乕儉丄偙傟偐傜壗偵廤拞偟偰傗偭偰偄偔偐丠丂僲僢僋傾僂僩偐丠丂偦傟偲傕亅亅亅亅丅丂枅擔丄媍榑傪偟偰歑偭偰偄傞丅

